


INTRODUCTION
Although the implementation of a universal hepatitis B vaccination programme has proven successful in preventing acute hepatitis B virus (HBV) infection [Reference Wen1], persistent infection with HBV remains a major global public health problem. There are about 240 million hepatitis B surface antigen (HBsAg) carriers worldwide [2] who are in the risk of acute and chronic liver diseases including cirrhosis and hepatocellular carcinoma (HCC) [Reference Chen3].
HBsAg is an important serological marker for the diagnosis and monitoring of chronic hepatitis B patients [Reference Cheng4]. HBsAg seroclearance during the clinical course can be considered as resolved hepatitis B infection [Reference Sorrell5]. HBsAg seroclearance may occur during the natural history of HBV infection and following antiviral therapy [Reference Lok6].
The incidence of spontaneous HBsAg seroclearance, defined as the absence of serum HBsAg on two occasions at least 6 months apart and remaining negative up to the last visit [Reference Liaw7], varies with geographical area. It is estimated that the annual incidence is about 0·7–1·7% in regions where the prevalence of HBsAg is low, such as Western countries [Reference Sampliner8–Reference Da Silva10], while that in regions where HBV is endemic, such as Asian countries, is about 0·5–3·08% [Reference Liaw7, Reference Chu11, Reference Han12].
Spontaneous HBsAg seroclearance is a rare event in chronic hepatitis B. However, most patients with HBsAg seroclearance have an excellent outcome and are in a state closest to a ‘cure’ [Reference Chu13], although some of them remain at risk of HCC [Reference Kim14]. Therefore, much effort has been expended to identify factors associated with HBsAg seroclearance with the objective of increasing the rate of HBsAg seroclearance. Older age, HBeAg seronegativity, clinical remission and cirrhosis have been reported to be associated with HBsAg seroclearance [Reference Chu13]. It also has been reported that HBV viral load and quantitative serum HBsAg levels can accurately predict HBsAg seroclearance [Reference Liu15]. However, little is known about the reappearance of HBsAg after spontaneous HBsAg seroclearance, which may constitute not only an increased risk of liver disease but also an infectious source.
In this study, based on our long-term cohort study [Reference Fang16], we report some asymptomatic HBsAg carriers who experienced spontaneous HBsAg seroclearance, with HBsAg reappearance many years later. We also determined whether the reappearance was caused by reinfection or reactivation using sequence analysis.
MATERIALS AND METHODS
Study subjects
All of the study subjects were recruited from the Long An cohort, which has been described previously [Reference Fang16]. The cohort was established in early 2004 from 27 340 agricultural workers aged 30–55 years living in the rural area of Long An county, Guangxi, China, using stratified sampling. It consists of 2998 HBsAg carriers. We followed the study subjects once every 6 months from 1 July 2004 to 1 July 2007. Then, we continued to follow-up some of the study subjects randomly once in 2013 when we randomly recruited 300 subjects with wild type of HBV core promoter and 300 subjects with HBV core promoter double mutations (1762T1764A) from the Long An cohort with the purpose of searching for human genes related to the double mutations. Each study subject provided a serum sample for the assessment of virological parameters and alpha fetoprotein concentrations.
Informed consent in writing was obtained from each individual. The study protocol conforms to the ethical guidelines of the 1975 Declaration of Helsinki and was approved by the Guangxi Institutional Review Board.
Serological testing
Sera were tested for HBsAg, HBeAg/anti-HBe, and anti-HBc using enzyme immunoassays (Beijing Wantai Biological Pharmacy Enterprise Co. Ltd, China). Alanine aminotransferase (ALT) levels were determined using a Reitman kit (Shanghai Fortune Long March Medical Science, China).
Measurement of viral loads
Serum HBV DNA concentrations were quantified by real-time polymerase chain reaction (PCR) using commercial reagents (Sansure Biotech Inc., China) in an ABI Prism 7500 sequence detection system (Applied Biosystems, USA) with a dynamic range of 1×102 –5 × 109 IU/ml.
Nested PCR for HBV DNA and nucleotide sequencing
DNA was extracted from 85 µl serum by pronase digestion followed by phenol/chloroform extraction. The first round of PCR was carried out in a 50 µl reaction using primers MD14 (nt 418–433, 5'-GCGCTGCAGCTATGCCTCATCTTC-3') and HCO2 (nt 761–776, 5'-GCGAAGCTTGCTGTACAGACTTGG-3'), with 5 min hot start followed by 35 cycles of 94 °C for 45 s, 45 °C for 45 s, and 72 °C for 60 s. The second round of PCR was carried out on 5 µl of the first-round products in a 50 µl reaction using primers ME15 (nt 455–470, 5'- GCGCTGCAGCAAGGTATGTTGCCCG-3') and HDO3 (nt 734–748, 5’-GCGAAGCTTCATCATCCATATAGC-3’), with 5 min hot start followed by 30 cycles of 94 °C for 45 s, 55 °C for 45 s, and 72 °C for 60 s. Products from the second round were confirmed by agarose gel electrophoresis. HBV DNA-positive products were sent to Sangon Biotech (China) for cloning sequencing. Amplicons were cloned into the vector pUCm-T (Sangon Biotech). Plasmid DNA was extracted using a SK1191 UNIQ-10 kit (Sangon Biotech) and the purified DNA was sequenced using a BigDye Terminator v. 3.1 Cycle Sequencing kit (Applied Biosystems) with sequencing primer ME15.
HBV serotyping
Serotypes were determined according to a single amino acid at the following positions: adrq- (122 K+127P+134 F+159 V+160R+177 A+178P), adrq+ (122 K+127P+134 F+159A+160R+177 V+178P), adw2 (122 K+127P+134 F+159A+160 K+177 V+178P), adwq+ (122 K+127 L+134 F+159A+160 K+177 V+178P), ayr (122R+127P+134 F+159A+160R+177 V+178P), ayw1 (122R+127P+134 F+159A+160 K+177 V+178P), ayw2 (122R+127P+134Y+159G+160 K+177 V+178P), ayw3 (122R+127 T+134 F+159G+160 K+177 V+178P) [Reference Courouce-Pauty17–Reference Chen19].
HBV genotyping
HBV genotypes were determined using phylogenies reconstructed on the basis of the complete S region (294 nt) of the viruses. The sequences were aligned to 16 HBV sequences of all known genotypes retrieved from GenBank using Clustal W and visually confirmed with the sequence editor BioEdit [Reference Hall20]. These sequences include M57663-Philippines, D23677-Japan, KT347089-South Africa, KM359441-Argentina, X75656-Polynesia, KF873543-Australia, KM999992-Philippines, EU670263-Philippines, Y07587-Germany, AB032431-Liberia, AY090456-Nicaragua, AF160501-USA, AY090460-USA, AF241407-Vietnam, AB486012-Japan and AY226578-Woolly monkey. Neighbour-Joining trees were reconstructed under the Kimura two-parameter substitution model with the program MEGA [Reference Tamura21]. The reliability of clusters was evaluated using interior branch test with 1000 replicates, and the internal nodes with over 95% support were considered reliable.
Statistical analysis
Statistical comparison of variables was performed using the χ 2 test. The 95% confidence intervals (CI) for the prevalence of variables were estimated. Logistic regression analysis was carried out to identify factors associated with spontaneous HBsAg seroclearance. All P values were two-tailed and P < 0·05 was considered to be significant. Statistical analysis was performed using SPSS v. 16.0 software (SPSS Inc., USA).
RESULTS
Baseline characteristics
In this study, we carried out seven rounds of sampling. Sampling of the study subjects depended on their consent. The seventh sampling, incorporated in another study, included a subset of the subjects in 2013. Again, the study subjects provided samples after giving consent. The total rate of HBsAg negativity was 2·0% (201/12355, 95% CI 1·4–1·8). The rate of HBsAg negativity in males (1·6%, 101/6413) was not significantly different from that in females (1·7%, 100/5942) (χ 2 = 0·225, P > 0·05). The rate of HBsAg negativity in the group originally HBeAg positive (0·3%, 5/1821) was significantly higher than that in the HBeAg-negative group (0·18%, 179/10134) (χ 2 = 22·668, P < 0·05). The difference in the rate of HBsAg negativity between the abnormal ALT at baseline group (1·4%, 23/1691) and normal ALT at baseline group (1·6%, 161/10264) was not significant (χ 2 = 0·416, P > 0·05) (Table 1). Clearly, HBeAg status is associated with HBsAg negativity.
Table 1. Results of the follow up of the cohort
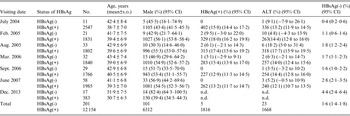
CI, Confidence interval; ALT, alanine aminotransferase; n.d., not done.
Spontaneous HBsAg seroclearance and its factors in the first six samples
According to the definition of spontaneous HBsAg seroclearance [Reference Liaw7], we found that this occurred in 41 subjects during 7583 person-years of follow-up evaluation, giving a 0·54% annual seroclearance rate. Multivariable logistic regression analysis was performed to identify factors associated with spontaneous HBsAg seroclearance. The independent variables included gender, age, genotype, viral load, HBeAg and ALT. Information for these variables was based on the baseline data. The results showed that older age (> 40 years) was associated with spontaneous HBsAg seroclearance [odds ratio (OR) 6·0, 95% CI 1·3–21·4, P = 0·022]. Low viral load (<103 copies/ml in serum) was also a factor for spontaneous HBsAg seroclearance (OR 6·3, 95% CI 1·4–28·8, P = 0·018) (Table 2).
Table 2. Logistic regression analysis for factors associated with spontaneous HBsAg seroclearance

OR, Odds ratio; CI, confidence interval; ALT, alanine aminotransferase.
* B, regression coefficient.
Reappearance of HBsAg after spontaneous seroclearance
Thirteen of the 41 individuals who experienced spontaneous HBsAg seroclearance were included in another study and therefore tested for HBsAg in 2013. Surprisingly, we found that four of these subjects had become positive for HBsAg after spontaneous seroclearance more than 8 years previously, making the rate of HBsAg reappearance 30·8% (4/13). None of the 13 individuals had a risk factor for reactivation, such as immunosuppression, steroid use, HIV infection, etc. The four subjects were QS840, TF336, TN98 and WX227; subjects QS840, TF336 and TN98 were positive for anti-HBs when HBsAg reappeared. Subjects QS840 and WX227 had abnormal ALT levels. It is difficult to explain this phenomenon by sex, age, genotype, viral load, HBeAg and ALT (Table 3). In order to determine whether the reappearance of HBsAg is due to reinfection or reactivation, we randomly selected three of the four with HBsAg reappearance for sequencing of the HBV genome.
Table 3. Characteristics of subjects with HBsAg seroclearance and reappearance

ALT, Alanine aminotransferase; n.d., not done.
Reappearance of HBsAg is due to reactivation
For subject QS840, 11 and 12 clones were selected from serum collected in 2004 and 2013, respectively. Serotypes adrq+ and adw2 were identified in the sample from 2004 but only serotype adrq+ was found in the sample from 2013. Subgenotypes C2 and I1 were identified in the sample from 2004. However, only subgenotype C2 could also be seen in samples collected in 2013. Ten of the 12 sequences from the sample collected in 2013 have the same amino-acid substitution mutation pattern in the major hydrophilic region (T118 K, T123N, G145A) as the sample collected in 2004 (Table 4, Figs 1, 2). All of the sequences from samples collected in 2013 have a similarity rate of 99% to that of 2004 at the nucleotide level. Clearly, the virus in the 2013 samples come from the virus in 2004 because they share the same serotype adrq+, subgenotype C2, mutation pattern and high rate of nucleotide similarity. Therefore, these data suggest that the reappearance of HBsAg in subject QS840 is due to reactivation.
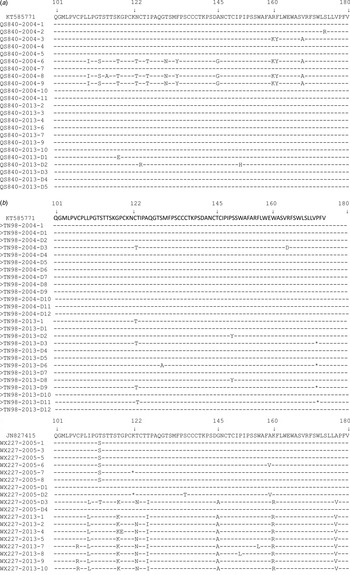
Fig. 1. Frequency and distribution of amino-acid substitutions in the MHR of HBsAg from each clone of (a) subject QS840, (b) subject TN98, (c) subject WX227.
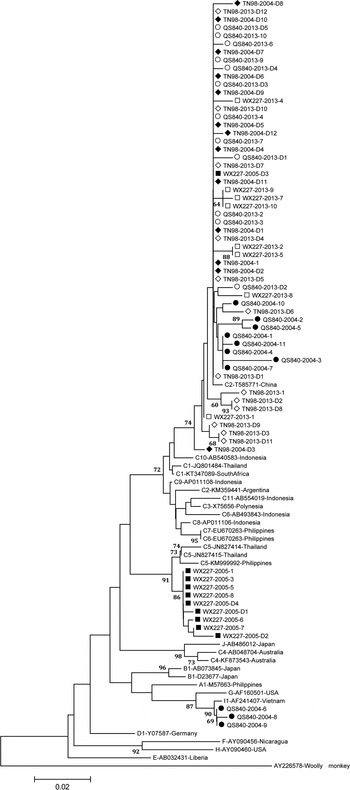
Fig. 2. Neighbour-joining trees. The trees were reconstructed on the basis of the S region (294 nt) of the viruses under the Kimura two-parameter substitution model with the program MEGA [Reference Tamura21]. The branch lengths represent the number of substitutions per site. The reliability of clusters was evaluated using the interior branch test with 1000 replicates and the internal nodes with over 75% support are considered reliable.
Table 4. Molecular characteristics of sequences from subject with reappearance of HBsAg
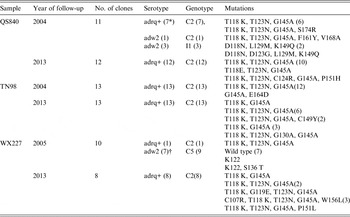
* Number of clones.
† Two sequences could not be serotyped because they have a stop codon at aa122.
For subject TN98, 13 and 13 clones were selected from serum collected in 2004 and 2013, respectively. Serotype adrq+ was the only serotype in both samples. Subgenotype C2 was also the only genotype in both samples. Most sequences from 2004 and 2013 share the same amino-acid substitution mutation pattern in the major hydrophilic region: T118 K, T123N, G145A (Table 4, Figs 1, 2). Five of the 13 DNA sequences from 2013 are the same as that from 2004. The remaining sequences have a similarity rate of 99% to that from 2004. Clearly, the reappearance of HBsAg in subject TN98 is also due to reactivation.
For subject WX227, 10 and eight clones were selected from serum collected in 2005 and 2013, respectively. There are two serotypes in the sample from 2005: adrq+ and adw2. However, serotype adrq+ was the only serotype in samples from 2013. There are two subgenotypes in samples from 2005: C2 and 5. However, only subgenotype C2 could be found in one clone in samples from 2013. Most sequences from 2005 were wild type while none of the sequences from 2013 were wild type in the major hydrophilic region (Table 4, Figs 1, 2). All DNA sequences from 2013 had a similarity rate of 99% to that from the only clone with subgenotype C2 from samples of 2005 and share the same mutation (T118 K and G145A). Their serotypes are all adrq+. These data also suggest that the reappearance of HBsAg in subject WX227 was due to reactivation.
Clearly, based on serotype, genotype, amino-acid substitution mutation pattern in the major hydrophilic region and similarity of nucleotide sequence in the S gene, the reappearance of HBsAg in this study is due to reactivation, suggesting that the definition of spontaneous HBsAg seroclearance [Reference Liaw7] has been challenged. Interestingly, all sequences involved in HBsAg reappearance encoded a G145A substitution before HBsAg seroclearance. However, none of sequences without G145A in samples QS840 and WX227 appear after reactivation (Table 4).
DISCUSSION
This study, based on the Long An cohort, is the first to report that reappearance of HBsAg may occur in individuals with HBsAg spontaneous seroclearance many years previously. The reappearance resulted from reactivation, rather than reinfection. We also found that the rate of spontaneous HBsAg seroclearance varies with cohort, not geographical area. Older age (>40 years) and low viral load (<103copies/ml in serum) were associated with spontaneous HBsAg seroclearance. However, factors that explain the reappearance of HBsAg remain unclear. The strength of this study is that it was based on a long-term followed-up cohort, which allowed us to obtain information from asymptomatic HBsAg carriers to spontaneous HBsAg seroclearance, and then HBsAg reappearance many years later. Furthermore, none of our study subjects were treated with antiviral drugs, which should avoid the influence of these drugs on HBsAg seroclearance and reappearance. The weakness of this study is that we did not test all of incidences of spontaneous HBsAg seroclearance, which may provide more information about reappearance and make the conclusion stronger.
Although HBsAg seroclearance was considered to be the state closest to a ‘cure’ [Reference Chu13], reactivation after seroclearance remains possible. This phenomenon is not a rare event in individuals with HBsAg seroclearance through antiviral therapy [Reference Perrillo22, Reference Hsu23]. It could also be observed in people with other diseases, such as HCC [Reference Jang24], haematological malignancies [Reference Han25] and HIV infection [Reference Waite26, Reference Vento27]. However, HBsAg reappearance in the general population with spontaneous HBsAg seroclearance has not been reported. Furthermore, none of the studies determined whether reinfection or reactivation was responsible for the reappearance of HBsAg. In this study, we are the first to report that HBsAg reappearance occurred in the general population and found that reactivation was the cause of its reappearance.
The natural course of chronic HBV infection can be divided into four phases, based on the virus–host interaction: immune tolerance, immunoreactive, low or no replication, and reactivation. The third phase is characterized by undetectable or low levels of HBV DNA [Reference Fattovich28]. In general, serum HBsAg titre has a positive correlation with HBV DNA concentrations [Reference Deguchi29]. It has been reported that patients with HBsAg seroclearance still harbour HBV within the liver (mainly in the form of cccDNA) up to 4 years after seroclearance, albeit at a very low replicative level and in a transcriptionally inactive phase [Reference Chu13]. It is possible that HBV exists within the liver for longer and causes reactivation when the host becomes disadvantaged, such as through immune suppression. If so, this could explain not only the reappearance of HBsAg but also why those experiencing HBsAg seroclearance remain at risk for HCC [Reference Kim14]. Therefore, our findings are important to remind clinicians to continue to monitor cirrhosis and HCC for individuals with HBsAg seroclearance, because spontaneous HBsAg seroclearance during the course of HBV infection does not mean resolved hepatitis B infection. It is also important for medical workers to monitor HBsAg because its reappearance constitutes a potential source of infection.
Generally, the incidence of spontaneous HBsAg seroclearance is lower in low-endemic areas such as Western countries than in high-endemic areas [Reference Ferreira30]. Recent findings suggest that the cumulative rate of HBsAg seroclearance in asymptomatic adult carriers from high-endemic areas is higher [Reference Chu11]. In this study, we found that the annual seroclearance rate is lower than that observed in low-endemic areas (European Caucasian subjects, 0·54% vs. 1·43%) [Reference Lauret31] and high-endemic areas (Taiwanese subjects 0·54% vs. 1·15%) [Reference Chu11], suggesting that the rate of spontaneous HBsAg seroclearance varies with cohort, not geographical area. We also found that older age and low viral load were associated with spontaneous HBsAg seroclearance, which is in accordance with other studies [Reference Ferreira30].
Although we found that reappearance of HBsAg was caused by reactivation, we could not reliably identify factors associated with reactivation. In this study, we found serotype adrq+ and adw2 in subjects QS840 and WX227 before seroclearance. However, only serotype adrq+ was found in their samples after reappearance. Furthermore, before seroclearance, adw2 was the predominant serotype in sample WX227 and serotype adrq+ was only found in one of the ten clones. It seems that serotype adrq+ is associated with reactivation. Another interesting observation is that only sequences with the G145A substitution appeared after reappearance. There are other sequences without G145A in the samples from QS840 and WX227 before seroclearance but none of them are present after reactivation. G145A mutation is well known as an escape mutation [Reference Lazarevic32]. The G145A mutation may result in significantly decreased HBsAg expression [Reference Martin33]. When the G145A mutant become predominant, it may reduce the level of HBsAg below the limit of detection so the subjects with this mutant appear to have achieved seroclearance. When the G145A mutant is a minor population, the level of HBsAg is above the limit of detection. However, this hypothesis requires further confirmation.
Reinfection is a possible cause of reappearance of HBsAg. However, none of the three subjects tested in this study had experienced reinfection. This may be attributable to the small sample size. Although 41 individuals with HBsAg spontaneous seroclearance were found in this study, we only tested 13 of them.
In conclusion, HBsAg may reappear in individuals with spontaneous HBsAg seroclearance many years previously. Reactivation is a cause of the reappearance. Therefore, it seems necessary to maintain clinical monitoring and regular surveillance for HBsAg and liver diseases after spontaneous HBsAg seroclearance.
ACKNOWLEDGEMENTS
We are indebted to staff members of Centre for Disease Prevention and Control of Long An and local town hospitals in Long An county, Guangxi, who assisted in recruiting the study subjects and sample collection, and to staff members of the Department of Virology, Centre for Disease Prevention and Control of Guangxi, China for their help in recruiting the study subjects and handling the sera. This study was supported by the Wellcome Trust (WT072058MA) and High Level Academic Backbone 139 Program in Medical Science of Guangxi.
DECLARATION OF INTEREST
None.






