


This paper contains supplementary material that can be found online at http://journals.cambridge.org
Introduction
Habitat fragmentation and loss, and degradation of habitat quality from human activities are among the main threats to wildlife populations (Wilcox & Murphy, Reference Wilcox and Murphy1985; Andren, Reference Andren1994; Fahrig, Reference Fahrig2003; Mortelliti et al., Reference Mortelliti, Amori and Boitani2010a). Fragmentation processes can also lead to population extinction in small areas (Debinski & Holt, Reference Debinski and Holt2000; Fischer & Lindenmayer, Reference Fischer and Lindenmayer2007), and recolonization processes become critical for specialist species that use a limited number of habitat types in the landscape (Franken & Hik, Reference Franken and Hik2004).
Large and long-lived mammals are generally recognized to be highly sensitive to human pressures and habitat change. Studies on large mammals are mostly limited to solitary species (Mortelliti et al., Reference Mortelliti, Amori, Capizzi, Rondinini and Boitani2010b) and only rarely focus on species living in social groups (Graham et al., Reference Graham, Douglas-Hamilton, Adams and Lee2009; Hewison et al., Reference Hewison, Vincent, Joachim, Angibault, Cargnelutti and Cibien2001). Social organization (group size, degree of philopatry of females, sex-biased dispersal, spatio-temporal stability of home ranges) is an important determinant of the ability of a social species to survive in a disturbed landscape and to colonize habitat fragments. In particular, habitat specialists and highly social species living in large groups are among the most negatively affected by habitat fragmentation and degradation (Swihart et al., Reference Swihart, Gehring, Kolozsvary and Nupp2003). Sociality and sex-biased dispersal may impose limits on colonization rates or on the size of a patch that can support a viable population because social groups are key parameters to population growth. It is still unknown to what extent patch quality and landscape structure (patch size, patch shape, isolation) affect the persistence of large forest-dependent mammals living in social groups.
To address these questions, we studied a forest-dependent and long-lived social species, the Barbary macaque Macaca sylvanus. This species lives in large multi male–multi female social groups of 10–88 individuals (Ménard & Vallet, Reference Ménard and Vallet1993a). Females are philopatric, living all their life in their natal group, whereas the males disperse. The wild Barbary macaque's distribution is limited to scattered and isolated forest fragments in Algeria and Morocco, most of them containing < 500 individuals. The largest population is in the Middle Atlas of Morocco, assumed to contain c. 75% of the north African population (c. 15,000 individuals; Taub, Reference Taub1977; Fa, Reference Fa1984). Decline of Barbary macaques in the Middle Atlas during the last 20 years has been attributed to the loss of cedar forests (Camperio Ciani et al., Reference Camperio Ciani, Palentini, Arahou, Martinoli, Capiluppi and Mouna2005) and the taking of young Barbary macaques for the pet trade (van Lavieren, Reference van Lavieren2008; van Lavieren & Wich, Reference van Lavieren and Wich2009). The remaining forest fragments of the Middle Atlas suffer from heavy human pressure, overgrazing by sheep and goats, and cedar pruning during shortages of food for livestock (Lamb et al., Reference Lamb, Damblon and Maxted1991). Barbary macaques forage > 50% of their time on herbaceous and shrub plants (Ménard & Vallet, Reference Ménard, Vallet, Fa and Lindburg1996). Therefore, domestic livestock are strong competitors. The Barbary macaque is categorized as Endangered on the Red List of Threatened Species (Butynski et al., Reference Butynski, Cortes, Waters, Fa, Hobbelink and van Lavieren2008) and is listed in Appendix II of CITES (CITES, 2013).
Our study aimed to provide up-to-date information on the distribution and abundance of the Barbary macaque in the Middle Atlas of Morocco. We compared the influence of landscape structure (patch area and shape, and patch connectivity), and habitat composition and quality, on macaque density. We considered the intensity of forest use by humans for livestock as an index of habitat quality, and also potential effects of altitude, which could interact with human pressure in montane regions.
Study area
The study area comprised 985 km2, mainly in the Ifrane National Park (Middle Atlas, Fig. 1), and included all areas subject to management by foresters. The Middle Atlas forest is mainly composed of mixed evergreen cedar–oak forests (Cedrus atlantica and Quercus rotundifolia). The study area is divided into numbered parcels by foresters, who carry out a regular silvicultural management programme consisting of cedar logging and the clear-cutting of holm oaks.

Fig. 1 The study area, showing the 14 forest patches potentially suitable for the Barbary macaque Macaca sylvanus, and unsuitable forest patches, and the locations where we surveyed for the macaque using line transects and complete counts. White shading indicates cultivated areas and villages. The areas where we carried out complete counts at Sidi M'Guild North, Azrou and Seheb were 17.1, 29.11 and 15.21 km2, respectively. The shaded area on the inset (Ifrane National Park) indicates the location of the main map in the Middle Atlas of Morocco.
Methods
Characterization of habitat types and degree of habitat fragmentation
We drew up a vegetation map from Sogreah-Ttoba (Reference Sogreah-Ttoba2004), which was updated with recent vegetation changes. We then classified each vegetation parcel as suitable or unsuitable for Barbary macaques, based on knowledge of their ecology (Fig. 1). Each Barbary macaque group uses a relatively stable home range. Barbary macaques need tall mature trees to avoid predators, either for nocturnal sleeping sites or to escape from danger during their daytime foraging activities. Consequently, open grasslands, immature oak coppices, degraded forest with heavily pruned or scattered cedars, and agricultural areas constitute an inhospitable matrix that macaque groups have never been observed to cross (Taub, Reference Taub1977; Ménard & Vallet, Reference Ménard, Vallet, Fa and Lindburg1996, Reference Ménard and Vallet1997).
A total of 14 forest fragments whose size exceeded the home range (3–4 km2) of Barbary macaque groups (Ménard & Vallet, Reference Ménard, Vallet, Fa and Lindburg1996) were retained as sampling plots (Fig. 1). We described each forest patch using seven habitat descriptors: vegetation community, index of human pressure, mean altitude, area, shape, and two measures of patch connectivity (Kindlmann & Burel, Reference Kindlmann and Burel2008; Prugh, Reference Prugh2009), here referred to as distance and connectivity. The index of human pressure was based on estimations of the intensity of overgrazing by sheep and goats and the intensity of cedar pruning in each forest patch (Sogreah-Ttoba, Reference Sogreah-Ttoba2004; see Supplementary Table S1 for calculations). Shape index was calculated using Eq. (1), where p i is the perimeter of the patch i and a i is its area (McGarigal et al., Reference McGarigal, Cushman, Neel and Ene2002).
 $${\rm Shape} = \displaystyle{{\,p_i} \over {2\sqrt {\pi ^*a_i}}} $$
$${\rm Shape} = \displaystyle{{\,p_i} \over {2\sqrt {\pi ^*a_i}}} $$
Distance, Eq. (2), is the minimum distance to the nearest occupied patch, where h ij is the nearest edge-to-edge distance between the focal forest patch and its neighbouring forest patch.
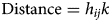 $${\rm Distance} = h_{ij} k$$
$${\rm Distance} = h_{ij} k$$
Connectivity is a distance-weighted area of the occupied patch, using Eq. (3), where d ij is the distance (km) between focal patch i and patch j, A j is the area (km2) of the forest patch j, and α is 1/drl. As social groups are units of colonization in this species (see Discussion), drl is the maximum observed day range length for a Barbary macaque group (3.00 km; Ménard & Vallet, Reference Ménard and Vallet1997), which allows a group to cross the matrix from one sleeping site to another within 1 day.
 $${\rm Connectivity} = \sum\limits_{\,j \ne i} {\exp ( - {\rm \alpha} d_{ij} )A_j k} $$
$${\rm Connectivity} = \sum\limits_{\,j \ne i} {\exp ( - {\rm \alpha} d_{ij} )A_j k} $$
Census methods
Censuses were conducted in the 14 forest fragments. Variables related to censuses were spatially referenced using ArcView v. 9.2 (ESRI, Redlands, USA). When possible we preferred to use complete counts to estimate densities but this method requires intense sampling that was impracticable over large areas (Fashing & Cords, Reference Fashing and Cords2000; Marshall et al., Reference Marshall, Lovett and White2008). Consequently, we used the line-transect method in several patches. At least 40 observations are needed to estimate densities from line-transect surveys (Marshall et al., Reference Marshall, Lovett and White2008) and we therefore applied a correction factor to density estimates obtained in forest patches where line transects alone were used, after determining the relationship between densities derived from complete counts and line transects in forest patches where both methods were used. We used both methods in five forest patches, only line transects in three patches and only complete counts in six patches (Fig. 1).
Censuses using line transects were carried out in October 2007 and 2008. October is the most suitable month for such surveys because the diurnal activity of the macaque is relatively stable at this time (Ménard & Vallet, Reference Ménard and Vallet1997), thus avoiding sampling biases from variations in the probability of detection. We sampled 10 line transects in eight forest patches comprising 526 km2 (Fig. 1). Each transect was 6 km (four comprised two lines of 3 km) and was repeated at least four times, except for one that was repeated three times. The total distance surveyed was 234 km. The beginning and direction of the lines were chosen to ensure that the surveys were entirely within forest patches.
There were four survey teams, each of two people. Censuses started at 06.00. Speed along transects was a maximum of 1 km h−1. At each sighting of macaques the observers noted the date, hour, location (with a global positioning system), and distance and angle to the first macaque sighted. The first sighted macaque was assumed to be located at the edge of the group. As groups have a greater chance of being detected than individuals (Buckland et al., Reference Buckland, Plumptre, Thomas and Rexstad2010), macaque groups were considered the sample units. The centres of detected groups were subsequently located on the map based on the estimated spread of the group (c. 100 m for a group of 15–25 individuals and c. 150 m for a group of 25–40; authors, unpubl. data) and the perpendicular distances from each group centre to transect line were estimated using ArcView.
Complete counts were carried out in 11 forest patches between March 2007 and October 2008, in sampled areas in three sites and over the whole of eight sites (Fig. 1). Observers made accurate counts of entire groups as they crossed a road or a track. The description of several individuals in each group allowed its identification.
Estimation of densities
Perpendicular modelling of group centres uses a detection function of perpendicular distances from a group centre to the transect line. We used DISTANCE 6 release 2 (Thomas et al., Reference Thomas, Buckland, Rexstad, Laake, Strindberg and Hedley2010) to fit a set of detection probability models to the observed perpendicular distances. We selected the best model according to the smallest value of Akaike's information criterion corrected for small samples (AICc; Burnham & Anderson, Reference Burnham and Anderson2002). In this manner, DISTANCE estimates the proportion of groups missed by the survey and computes the population density and size in the survey area. A mean group size in each forest zone sampled was implemented as a parameter for density estimation with DISTANCE (Table 1).
Table 1 Densities (D s, number of groups km−2; D, number of individuals km−2), encounter rates of groups (n/L; n, number of observations of groups; L, total survey effort in km), and abundance of the Barbary macaque Macaca sylvanus in the 14 forest patches (the data for the patches Bekrit, Ras el Ma and Senoual were pooled) surveyed in the Middle Atlas (Fig. 1), estimated from complete counts and line-transect surveys.

1 Mean group sizes(s) obtained from complete counts of several groups (n) were used in DISTANCE for density calculation (see text for further details) at Sidi M'Guild North (s = 33, n = 12), Azrou (s = 17, n = 12), Seheb (s = 20, n = 8), Bekrit (s = 27, n = 11), Senoual (s = 23, n = 3) and Ras el Ma (s = 17, n = 3).
2 Coefficient of variation of density
3 Both line transects and complete counts were conducted. Within the areas sampled in Azrou, Seheb and Sidi M'Guild North (Fig. 1) densities (D s and D) were estimated from complete counts, including 207 individuals (twelve groups), 171 individuals (eight groups) and 400 individuals (12 groups), respectively.
4 Only line transects were used. The three sites (Bekrit, Ras el Ma, Senoual) where few macaque groups were detected during line transects were grouped for density analysis using DISTANCE.
5 Densities obtained by line transects for the three pooled patches were corrected by dividing by 1.6, and the new values were considered reliable estimates of those that would have been obtained by complete counts (see text for further details).
6 We did not obtain exact group sizes because groups did not cross paths.
7 The sites where no macaques were detected during transects were excluded from analyses with DISTANCE.
8 Only complete counts were conducted.
Influence of the environment on density
We examined the influence of four descriptors of the vegetation community (% cedar, cedar–oak or pine forest, % mature oak forest, % oak coppice, % open grassland or shrubby formation) on the density of individuals and groups of macaques. Data analysis was in two stages. Firstly, we computed a principal component analysis (PCA) of the environmental descriptors (four variables, 11 forest patches); three of the 14 patches were excluded because they were pooled for density estimation (see Results). Data were arcsin-transformed before analysis. We then carried out a regression of macaque abundance against the first two axes of the PCA. All analyses were carried out with R (R Development Core Team, 2010).
A multiple linear regression (function lm in R) was used to examine the effect of six environmental variables (human pressure, altitude, area, shape, distance and connectivity; Supplementary Table S1) on macaque density. Model selection procedure was based on AICc. The model with the lowest AICc was chosen as the best fitting and the most parsimonious. Models with ΔAICc < 2 from the best model were considered equally effective for explaining variations in density. For each model we calculated AIC weight (w i ), which gives the relative importance of each variable in explaining variations in density. These calculations were made using the MuMIn package in R.
Results
Vegetation types and forest patches
The study area is a complex vegetation mosaic. Pure and mixed mature forests cover c. 58% of the study area, oak coppices 22%, and open grasslands and scattered shrubby formations 20%. A total of 539.2 km2 was judged suitable for macaques (54%, Fig. 1). The inhospitable matrix (agricultural areas excluded) was mainly composed of open vegetation, grasslands or shrubby formations (26%), low and scarce oak coppices (36%) and sparse and pruned cedars (28%). Each forest patch was composed of a mosaic of vegetation formations (Supplementary Table S2). Eleven forest patches were mainly mixed cedar–oak formations (> 50%) and two patches were mostly pure oak formations. Each forest patch contained 2–23% open grassland or shrubby formations scattered in small areas. The area of forest patches was 5–142 km2.
Macaque density
Estimated macaque densities were 0–23.4 individuals per km2 and 0–0.7 groups per km2 (Table 1, Supplementary Fig. S1). Seven forest patches each contained < 100 individuals. We detected 40 groups with line transect surveys. Calculations in DISTANCE showed that the uniform+cosine model had the lowest AICc value. The half strip width within which macaque groups had the highest probability (100%) of being detected was 161.9 m. This value exceeded the distance at which macaques were detected (mean 39 m; range 10–70 m) because group centres were used in analyses rather than the position of the first macaque sighted. The results obtained from complete counts and line transect surveys varied comparably between sites. However, densities were overestimated by line transects, by a factor of 1.4–1.8, compared to densities obtained from complete counts at the same sites (Table 1). We used densities from line transects, divided by a correction factor of 1.6, to estimate densities at the three sites (Ras El Ma, Bekrit, Senoual) where we only surveyed with line transects (Table 1). The total number of macaques in the sampled area was estimated to be 4,972. The mean density among the sampled forest patches was 9.2 individuals per km2.
Influence of environmental variables on density
The PCA of the four vegetation descriptors clearly separated the 11 forest patches (data not presented). Axis 1 accounted for 65.6% of the variation in the data and distinguished forest patches dominated mostly by oak formations (mature oak forests or oak coppices; e.g. Aïn Leuh) from those occupied by mixed cedar–oak or pine forest (e.g. Nokra, Seheb), and Axis 2 for 21.8%, grouping patches according to the percentage cover of open areas such as grasslands, shrubby formations and oak coppices (e.g. Azrou, Jbel Ben IJ). Macaque density did not have any relationship with the PCA coordinates of forest patches, indicating that the abundance of macaques was not sensitive to the characteristics of forest patch vegetation (Fig. 2).

Fig. 2 Linear regression between macaque density and (a) Axis 1 (PC1) and (b) Axis 2 (PC2) of a principal component analysis (PCA) of the vegetation descriptors of the 11 forest patches (Table 1).
Multiple linear regressions with six environmental variables as predictors of macaque density, in 11 forest patches, indicated that human pressure was the best predictor (Table 2). The second most supported model (ΔAICc < 2) included only area (Table 2). The sum of all Akaike weights containing human pressure indicated that it was the most important variable. Densities significantly decreased as human pressure increased (F 1,9 = 8.39, P = 0.018, R 2 adj = 0.43) and increased when patch area increased (F 1,9 = 5.85, P = 0.038, R 2 adj = 0.33, Fig. 3a,b). However, when we excluded the largest patch the correlation between density and patch size was not significant (F 1,8 = 0.06, P = 0.812, R 2 adj = 0.11). We found no relation between density and the shape or degree of connectivity of forest patches, or their altitude (F 1,9 < 1, P > 0.1).

Fig. 3 Linear regressions between macaque density and (a) an index of human pressure and (b) area of the 11 forest patches (Table 1).
Table 2 Model selection of predictors of macaque density based on Akaike's information criterion corrected for small sample size (AICc), and relative importance of predictor variables assessed by summing the AIC weights (w i ) from each model containing that predictor.

1 Patch shape, connectivity and area were not tested together because the calculation of each of them included a measure of area and they were therefore considered to be strongly correlated with each other. Distance and connectivity were not tested together because the calculation of each of them included a measure of distance. Human pressure and connectivity were not tested together because they were significantly negatively correlated (F 1,9 = 13.03, P = 0.005, R 2 adj = 0.55). The other predictors were independent (P > 0.05). Shapiro–Wilk normality test on the models did not show any departure from normality (P > 0.1).
2 Models with ΔAICc < 2 are considered equally plausible in explaining variations in density.
Discussion
The forested areas in the study site in the Middle Atlas of Morocco are partly transformed into oak coppices, a result of clear-cutting of holm oaks. Open areas of oak coppices are unsuitable habitats for macaques, whereas pure mature oak forests may harbour at least 20 km−2 (Fa, Reference Fa1984). With recent climate change, which has led to a decrease in winter snow, some shepherds stay in the high mountains during the winter, and during food shortages they prune cedars to feed their livestock (Lamb et al., Reference Lamb, Damblon and Maxted1991). These practices, combined with overgrazing by sheep and goats, have led to a gradual disappearance of forests, soil erosion, and loss of diversity and abundance of herbaceous and shrub resources in comparison with preserved cedar–oak forests (Ménard & Qarro, Reference Ménard and Qarro1999). The consequence of habitat loss and degradation of forest areas is fragmentation of habitat suitable for macaques into forest patches of varying size.
Comparisons with previous censuses at two sites in the Central Region of the Middle Atlas (Azrou: 10 individuals km−2; Ain Kahla, i.e. Sidi M'Guild North forest: 36–44 individuals km−2; Taub, Reference Taub1977; Fa, Reference Fa1984) vs 7.1 and 23 individuals km−2 in our study show a decrease in density of c. 40% in 30 years. This is in agreement with a study carried out in 2005 in the same two forests (van Lavieren & Wich, Reference van Lavieren and Wich2009). However, van Lavieren & Wich (Reference van Lavieren and Wich2009) observed smaller mean group sizes (6–9) compared with those that we found with our complete counts (17–33), and higher mean group density (2.1–3.3 km−2 vs 0.6–1.1 km−2), which suggests they observed several parties of groups that they considered to be different groups, perhaps because they were spread in the forest. Our estimate of a total population of c. 5,000 Barbary macaques in the Middle Atlas is about three times less than that estimated 30 years ago (c. 15,000; Taub, Reference Taub1977). The decline is probably a result of the decrease in macaque density in areas of suitable habitat and the disappearance of entire forest fragments. We found that macaque densities varied greatly between forest patches (0–23 km−2). A high intensity of human pressure and/or small patch size appear to be responsible for the low macaque densities. However, the lack of relationship between density and patch size when the largest patch of 140 km2 was removed from the analysis suggests that a possible area threshold exists between this largest patch and the second largest of c. 80 km2, below which the size effect disappears. Macaques may depend more on the quality of herb and shrub layers, related to the intensity of livestock grazing, than on forest structure. We did not find evidence of an effect of patch connectivity, which suggests a lack of functional connectivity between patches and that patches may be too far apart for macaques to move between them.
The study of behavioural ecology has facilitated the understanding of the effects of changes in landscape structure on the abilities of populations to persist in or recolonize an area (Lima & Zollner, Reference Lima and Zollner1996; Lawes et al., Reference Lawes, Mealin and Piper2000; Kie et al., Reference Kie, Ager and Bowyer2005). Social organization can also limit the resilience of a species to the effects of habitat loss, degradation and fragmentation but, to our knowledge, few studies have taken into account the influence of sociality (Lawes et al., Reference Lawes, Mealin and Piper2000; Swihart et al., Reference Swihart, Gehring, Kolozsvary and Nupp2003; Graham et al., Reference Graham, Douglas-Hamilton, Adams and Lee2009). We hypothesize that specialist habitat-dependent species of large mammal living in large social groups in which only one sex disperses will be particularly vulnerable to habitat fragmentation and loss because sociality may restrict colonization. Migrating male Barbary macaques alone cannot successfully colonize or recolonize an area: only social groups can do so. Group fission is the only way for females to disperse but, as observed in similar forests in Algeria, this leads to the formation of new groups that remain in the same forest patch (Ménard & Vallet, Reference Ménard and Vallet1993b). Barbary macaques are able to move outside the forest to feed in adjacent open grasslands or oak coppices (Ménard & Vallet, Reference Ménard and Vallet1997). However, studies of the home range use by several focal groups have not observed macaques crossing wide open areas or moving more than c. 200 m from the forest edge (Taub, Reference Taub1977; Ménard & Vallet, Reference Ménard, Vallet, Fa and Lindburg1996). We believe that the social organization of Barbary macaques, with the total philopatry of females and their strong, stable residence in their home ranges, high sociality and large cohesive groups, reliance on mature forests, and requirement for groups of trees for an entire group to sleep or to escape from danger, limits the likelihood of movement between patches. This probably explains why this macaque is sensitive to the quality and size of forest patches whereas density does not seem to be influenced by the structure of the landscape. These traits appear to predispose the species to local extinction because fragmented populations are likely to be entirely isolated.
In the near future we suspect that stochastic events will extirpate the Barbary macaque in forest patches where their density is close to zero (Melbourne & Hastings, Reference Melbourne and Hastings2008; Traill et al., Reference Traill, Brook, Frankham and Bradshaw2010). These patches will not be recolonized, even if suitable, because of the low dispersal abilities of this macaque and the reduced connectivity of the landscape. Forest patches are separated by areas that were formerly wooded but are now degraded. Approximately 64 km2 (14%) of these areas could be restored in the medium term, the time necessary for oak coppices to reach maturity. Such restoration could create corridors between six of the 14 actual forest patches, which would reduce the number of patches to nine (N. Ménard, unpubl. data). In addition, expanded forest patches could exceed the area threshold to become more favourable for the conservation of the macaque. In contrast, c. 23 km2 within four of the existing forest patches are heavily degraded and becoming more unsuitable for the macaque as a result of cedar pruning. We estimate that the outcome of this process will fragment these patches, increasing the number of forest patches from 14 to 18 (N. Ménard, unpubl. data). If human pressures persist the population of Barbary macaques in the Middle Atlas will be in danger of extinction. In addition, it has been predicted that with climate change C. atlantica will disappear from the Middle Atlas but will persist in the south-west High Atlas (Cheddadi et al., Reference Cheddadi, Fady, Francois, Hajar, Suc and Huang2009). Consequently, we recommend the development of mature holm oak forests instead of using clear-cutting management techniques, and favour the expansion of new forests at higher altitudes. The Middle Atlas will otherwise become dominated by mobile and generalist species (Devictor et al., Reference Devictor, Julliard and Jiguet2008).
In conclusion the population of the Barbary macaque in the Middle Atlas continues to decline, and locally low densities result from habitat loss, unsuitable forestry practices and competition with livestock. We therefore recommend halting the clear-cutting of holm oaks and restriction of the number of sheep and goats within the forest. Poaching of young macaques for illegal trade is an additional threat to this already declining population (van Lavieren, Reference van Lavieren2008; van Lavieren & Wich, Reference van Lavieren and Wich2009), leading to a deficit of immature individuals.
Acknowledgements
Financial support for this study came from the contract Service Provincial des Eaux et Forêts d'Ifrane/University of Rennes 1, with N. Ménard and M. Qarro as the French and Moroccan scientific leaders, respectively. We thank all the team of the Ifrane National Park project, Z. Amhaouch, I. Bouziane, B. Elasri, N. Elrouat, L. Oukannou and Mr. A. Sayad (Association of the Friends of the National Park of Ifrane), who participated in the line-transect sampling in 2007 and 2008. This study was conducted in close partnership with the Haut Commissariat aux Eaux et Forêts et à la Lutte contre la Désertification and the Regional Director of Eaux et Forêts du Moyen Atlas at Meknes. We thank P. Le Gouar for helpful comments, J. Nabucet for help with geographical information system analysis, F. Cuzin for information on Barbary macaque groups on Jbel Ben IJ, and two anonymous reviewers for helpful comments. We are grateful to Helen Whitfield for revising the language of this article. This is a scientific production of UMR CNRS 6553, Caren-OSUR, University of Rennes 1.
Biographical sketches
Nelly Ménard and Dominique Vallet have been studying the ecology and demography of the Barbary macaque since 1982. Yann Rantier is a specialist in geographical information systems. Adrien Foulquier is a veterinarian. Mohamed Qarro has been studying pastoralism in the Middle Atlas for many years. Lahcen Chillasse is developing research on the ecology of birds and mammals in Morocco. Jean-Sébastien Pierre specializes in population dynamics, modelling and statistics. Alain Butet focuses on research in landscape ecology.






