


Forms of Persistence
Ever since living beings arose from non-living organic compounds on a primordial planet, more than 3.5 billion years ago, a multitude of organisms has unceasingly flourished by means of the reproduction of pre-existing organisms. Through reproduction, living beings generate other material systems that to some extent are of the same kind as themselves. The succession of generations through reproduction is an essential element of the continuity of life. Not surprisingly, the ability to reproduce is acknowledged as one of the most important properties to characterize living systems. But let’s step back and put reproduction in a wider context, the endurance of material systems.
Compared to material systems belonging to the domain of inanimate matter, such as rocks and minerals, living beings are material systems with a relatively modest degree of physical persistence: their existence depends on some capacity for ‘renewal’ through time. This renewal occurs at different levels of their organization and at different time scales with respect to the lifetime of the individual organism.
At the level of their most basic components, all living beings are subject to a considerable flow of matter through growth and metabolism, which has profound effects on their constitution at the molecular level. At each breath, about 1021 oxygen atoms, which for a while have been part of our body, abandon us in the form of CO2 and H2O molecules, while as many new oxygen atoms, in form of O2 molecules from the air, become part of us. These ‘new’ atoms have certainly belonged to countless other living beings – microorganisms, plants, dinosaurs, bugs and multitudes of past and present humans, including ourselves some time ago. There are so many atoms even in a small chunk of matter, that this kind of atomic promiscuity is a statistical certainty. It has been calculated that more than 90% of our atoms are replaced every year. At any time, our bodies take in new atoms from the air we breathe, the food we eat, and the liquids we drink. These atoms are incorporated into our cells and fuel the chemical processes that keep us alive. This happens in all living beings, although to a variable extent, depending on the species and the life stage. For example, there is very little molecular turnover in the resting spores of many bacteria, which can remain dormant for thousands of years.
At a higher level of their organization, all living beings renew whole parts of their body during life. Unicellular organisms can renew subcellular structures, and, more prominently, multicellular organisms can renew entire cells. The turnover rate of our cells (calculated for an adult human male of 170 centimetres and 70 kilograms) varies among tissues, from 0 (cells never replaced, like most brain neurons) to 210 billion cells per day (red blood cells), for a total of 330 billion cells per day, corresponding to about 80 g of mass. About 86% of these cells are blood cells, and most of the remainder belong to the wall of the stomach and the gut. Our red blood cells persist in circulation for only about 3–4 months in an adult individual, before being phagocytized by other circulating cells, so they must be continuously replenished through cell proliferation and differentiation. The epithelial cells of our intestine are renewed every 4–5 days, so we renew more than 30 g of our gut (about 40 billion cells) per day. This kind of renewal at the cellular level is even more conspicuous in case of injuries to the body that are repaired through healing or regeneration, as when a lizard loses its tail.
Finally, since living beings are mortal (a topic we will discuss in detail before the end of this chapter), there is a population-level renewal, through reproduction. The plants that cover the slopes of a mountain are probably not the same individuals that covered those slopes a thousand years ago. A pine tree, before dying and disappearing as an individual material system, can generate other pines, and the pine forest endures, at least for a while, certainly for much longer than the time the single pine tree persists. On the longest time scale, living systems compensate by means of reproduction for their limited capacity for individual persistence.
All these forms of renewal in living beings contrast with the apparently unchanging shape of the mountain on which our pine forest sits. On the time scale of the life of the forest, the mountain has largely maintained its constitution down to the single atoms, most of which are the same and have remained in the same spatial relationships, the latter affected only to a negligible degree by movements of blocks of rock along fault lines and the surface phenomena of erosion and transport.
An exploration of this third way of living renewal–the reproductive processes – is the subject of this book, while the objective of this chapter is to equip ourselves with some conceptual tools that will be needed along our journey. To help the reader manage the diversity of organisms we will mention in the following pages, a quick guide to the diversity of life is provided in Box 1.1, while some basic biological knowledge needed to appreciate the diversity of reproductive process can be found in the last section of this chapter.
Box 1.1 A Classification of Living Organisms
To help readers move through the variegated landscape of our broad-spectrum taxonomic treatment, we outline here a classification scheme that is as up to date as possible. For convenience of exposition, we have nevertheless saved a few old names that have disappeared from current classifications because the groups they identify have been acknowledged not to be monophyletic, that is, to include all and only the descendants of a common ancestor. For example, while vertebrates are confirmed as a monophyletic group, invertebrates are simply all animals that are not vertebrates. Other commonly used names that do not correspond to monophyletic taxa are protists (unicellular eukaryotes), polychaetes (annelids other than oligochaetes, the latter including earthworms and leeches), crustaceans (in the traditional sense that excludes insects), algae, bryophytes, pteridophytes and even reptiles in the usual sense that excludes birds.
The primary taxonomic division of living beings is between prokaryotes and eukaryotes. Prokaryotes, most of which are unicellular, include the true bacteria, within which are classified also the blue-green algae, and a diverse group known as the archaea. Unicellular organisms also account for a huge and diverse set of eukaryotes, only a few of which have close affinities to multicellular ones, such as Volvox or Chlamydomonas with the true plants, or diatoms with the brown algae. Most unicellular eukaryotes, and a few multicellular ones that are closely related to some of them, are informally called protists. Free-living aquatic photosynthesizing protists are often called algae, an informal group that includes also aquatic photosynthesizing multicellular organisms; based on their pigments, red, brown and green algae are distinguished.
The most familiar groups of eukaryotes are the land plants, the fungi and the animals.
The simplest kind of land plants (or embryophytes) are mosses and liverworts, traditionally grouped as bryophytes; they lack the vessels for the transport of sap that characterize the remaining land plants, therefore called vascular plants or tracheophytes. These comprise, in turn, horsetails and ferns, collectively called the pteridophytes, and a much larger group, the seed plants or spermatophytes. Some of the latter, the gymnosperms, including conifers, lack true flowers and produce naked ovules; their sister group are the flowering plants, or angiosperms. Here, ovules mature within the ovary, and thus the seeds are enclosed in a fruit.
Long classified with plants, but closer instead to animals, are the fungi. These include a modest number of unicellular forms (yeasts) and a huge diversity of multicellular organisms. Devoid of the pigments that characterize algae and other plants, fungi are mostly exploiters of decaying matter, or symbionts associated with the roots of plants, or parasites. The main fungal taxa are ascomycetes, basidiomycetes and zygomycetes.
Within animals (or metazoans), some 30 highest-level groups or phyla are currently accepted. Sponges are mainly marine, but a few live in freshwater. Other marine groups are the diaphanous ctenophores or comb-jellies, and nearly all cnidarians (sea anemones, corals, medusae). Many free-living members of the platyhelminths or flatworms are also marine, but the group includes freshwater forms such as planarians. Flatworms include also two large groups of parasites, the digeneans or flukes and the cestodes or tapeworms.
A generally elongated body articulated into segments is the most obvious feature of annelids. These include terrestrial or freshwater groups like earthworms and leeches, but the majority is represented by the marine polychaetes. Other animal groups are often described as minor, because they include only a handful of species or because these are small and inconspicuous, or for both reasons, but we will nonetheless encounter them in our pages because of peculiarities of their reproductive biology.
Huge groups of invertebrates are the nematodes and the arthropods. Nematodes or roundworms include very many parasites, including Ascaris, but also free-living species such as Caenorhabditis elegans, one of the most popular model organisms studied in the laboratory. Arthropods, unmistakable for their segmented body provided with an exoskeleton and more or less numerous pairs of articulated appendages, including sensory (antennae), feeding (maxillae, mandibles) and locomotory (legs), are the largest of all animal phyla. They include the chelicerates, with scorpions and pseudoscorpions, mites and spiders; the many-legged myriapods; and the crustaceans, mainly aquatic, but with a successful branch of terrestrial forms (woodlice or land isopods). Other than the familiar decapods (crabs, shrimps, lobsters), crustaceans include many other groups such as cladocerans (Daphnia). Based on molecular evidence, insects are classified today by most authors as a specialized lineage of crustaceans.
Very unusual and diverse are the shapes of the echinoderms (e.g. starfishes and sea urchins), and those of tunicates, the sessile ascidians and the planktonic thaliaceans. Despite the obvious morphological distance, these latter groups of marine animals are closely related to vertebrates.
Within vertebrates, the old class of fishes has been long abandoned, their content being now distributed between several major groups: we will deal with representatives of the cartilaginous fishes (sharks, rays), crossopterygians (coelacanth) and numerous bony fishes. Of amphibians, we will mention both urodels (salamanders) and anurans (frogs and toads). Of reptiles other than birds, we will see the lineages of turtles, squamates (lizards, snakes) and crocodiles.
Following a simplified but up-to-date classification, the mammals mentioned in the text belong to the following groups: monotremes (platypus and echidnas), marsupials (kangaroo, wallaby, opossum) and placentals, to which we belong.
What Is Reproduction?
Let’s adopt an informal common-sense concept of reproduction. In biology, we can define reproduction as the process by which new individuals are generated from pre-existing individuals. Implicitly, it is assumed that the ‘new individuals’ are materially derived from pre-existing ones, or parents. This concept of reproduction has deep roots in human history, as it emerged by obtaining increasingly detailed knowledge of the life cycles of the plants and animals most familiar to us, ourselves included. This simple definition, as trivial as it may seem, nonetheless allows us to leave out other possible forms of generation that are not found in living beings.
A long tradition that remained alive until the nineteenth century accepted the possibility of spontaneous generation, that is, the direct formation of living organisms (the simplest ones at least) from inanimate matter, such as mud or rotting organic material. It was William Harvey (1578–1657), who in his Exercitationes de generatione animalium of 1651 first recognized a fundamental divide between the world of living beings and the rest of nature, based on a principle of generation: ‘omnia ex ovo’, all living beings are born of an egg. Belief in spontaneous generation was progressively dismantled through the experimental work of scientists, among them Francesco Redi (1626–1697), who inflicted a mortal blow to the doctrine of spontaneous generation with his experiments designed to reveal whether ‘any exudate from a rotten corpse, or any filth of something putrefied, generates worms’. Redi observed indeed that ‘worms’ (actually, insect larvae) developed in sealed glass flasks containing rotting pieces of meat or little dead animals, but only if, before sealing them, access to these flasks and their content was allowed to big flies, blue, green or grey, similar to those into which the ‘worms’ eventually transformed. Further important contributions were provided by Lazzaro Spallanzani (1729–1799), Franz Schulze (1815–1873) and finally Louis Pasteur (1822–1895), who definitively demonstrated that spontaneous generation does not occur, not even in microorganisms.
Material systems that are not considered living organisms, or do not qualify as living cellular systems – for instance, viruses, prion proteins (prions) and transposable DNA sequences (transposons) – can reproduce in some sense, although differently from living organisms. An important difference with respect to organismal reproduction is that these systems can replicate without contributing constitutive matter to their descendants. In other words, the causal link between ‘parents’ and ‘offspring’ does not pass through the transformation of a part of the body of the parent into the offspring. For instance, prions are proteins with a special conformation, capable of inducing other molecules of the same protein, but with a different three-dimensional structure, to take their own conformation. In this way, prions start a chain reaction that leads to their multiplication in an infectious way. Some prions are associated with pathological conditions of the mammal nervous system – for instance, bovine spongiform encephalopathy (BSE), also known as mad cow disease. Another example is offered by retroviruses, such as human immunodeficiency virus (HIV). When infecting a cell, the virus induces it to produce copies of its genetic material and the molecules for its shell, the capsid. However, neither the genetic material nor the proteins of the capsid of the ‘offspring generation’ are derived from parental molecules. The parent virus is causally responsible for the production of a new viral unit, but does not contribute any of the matter of which it is made.
Apart from these other forms of reproduction, two different aspects of population renewal contribute to biological reproduction. One is that ‘new individuals’ are added to the set of existing ones. In this demographic aspect of reproduction, ‘new’ should be understood as a quantitative addition to the entities that already exist. The other is the production of ‘individuals that are new’, compared to the existing ones. In this innovative aspect of reproduction, ‘new’ should be understood as qualitatively different from the pre-existing entities. According to the demographic concept, focus is on replacing the individuals that inevitably perish, thus maintaining or possibly increasing the size of the population. In contrast, according to the innovative concept, focus is on the appearance of ‘something new under the sun’, thus allowing the population to change through the generations – that is, to evolve.
These two distinct aspects of reproduction are found together in some forms of reproduction, for instance, in the most common instances of sexual reproduction, but not in all. As we will see, some modes of reproduction lead to a renewal only in the demographic sense, while innovation, although eventually occurring in all organisms, is not necessarily associated with reproduction.
Asexual and Sexual Reproduction
Traditionally, a primary distinction is traced between asexual and sexual reproduction. This division, as the two terms suggest, is based on the characterization and involvement of the so-called sexual processes.
At the genetic level, sexual processes are biological processes through which new combinations of genetic material are created from distinct sources. These can occur through the union of genetic material of different provenance, as in the fusion of the gametes of two individuals, but also through the reassortment of genetic material of different origin, like the reshuffling of the two sets of chromosomes during meiosis (recombination) for producing gametes or spores. Novel genetic assets can be obtained also in other ways. One is horizontal gene transfer, where genetic material is moved from one individual to another that is not its offspring (and not even necessarily of the same species), to be incorporated in the latter’s genome. This sexual process occurs regularly in prokaryotes, but it is common among eukaryotes and between prokaryotes and eukaryotes as well. Sexual processes, sometimes dubbed simply sex, for short, are thus distinguished, in principle at least, from the processes of change of genetic information that occur in a single individual without the contribution of DNA from other sources, as in gene mutations. In the case of eukaryotes, for simplicity, in this book we will discuss the genetic aspects of reproduction at the level of the nuclear genome, overlooking the dynamics internal to the genomes of the organelles (mitochondria and plastids).
These phenomena of genetic reassortment may or may not be associated with reproduction. A classic example of sex dissociated from reproduction is offered by ciliates, a group of unicellular protists to which paramecia belong. Ciliates commonly practise a form of sex called conjugation. Two individuals (conjugants) unite temporarily, exchange genetic material through a sort of reciprocal fertilization, then separate again. The result of this exchange is a pair of independent individuals (ex-conjugants), genetically different from the conjugants. Note that the number of individuals has not changed through sex: two conjugants in, two ex-conjugants out.
Thus, in ciliates it is possible to have reproduction without sex, when a paramecium simply splits into two (binary fission, Chapter 3), or sex without reproduction, when two paramecia unite, exchange genetic material and then separate, both genetically renovated (conjugation).
Sexual processes, associated or not with reproduction, are found in virtually all major lineages of living beings. Note that in the scientific literature about the genetics of reproduction, ‘to have sex’ generally means ‘to carry out a sexual process’, not ‘to mate’. That a mating may not result in reproduction is probably quite obvious to everybody.
Reproduction can thus be qualified as sexual or asexual based on the involvement of potentially independent sexual processes. Accordingly, sexual or asexual are in principle attributes of reproduction, but not attributes that apply to a given organism, since many organisms can reproduce in both ways.
Asexual reproduction (in plants, also called vegetative reproduction) is a mode of uniparental reproduction (i.e. reproduction from a single parent) that does not involve sexual processes or the production of gametes, not even in derived or residual form. As a first approximation, asexual reproduction generates individuals genetically identical to each other and identical to the parent, thus forming what is termed a clone (Chapter 3), and for this reason it is also called clonal reproduction. However, asexual reproduction in some cases fails to produce perfectly clonal descendants, while sexual reproduction occasionally has a clonal outcome.
Asexual reproduction is the most common form of reproduction among unicellular organisms, prokaryotes and eukaryotes alike, but is also very common among multicellular organisms. For instance, many plants, for example most reeds, propagate by runners; many marine worms multiply by splitting into pieces; and some sea anemones generate new polyps through budding. In bacteria and some protists, this is the only mode of reproduction (obligate asexual reproduction). However, more often, asexual reproduction coexists with sexual reproduction, either as the only form of reproduction in a specific phase of an organism’s life cycle (e.g. in the polyp phase of cnidarians with a typical metagenetic cycle; Chapter 2), or as a reproductive option (facultative asexual reproduction) co-occurring with the sexual one (e.g. in many plants). In any case, the obligate asexual reproduction of certain organisms does not rule out the possibility of these having sex that is not associated with reproduction, as mentioned above for the paramecium, and this is very common in prokaryotes.
Sexual reproduction is a form of reproduction that generates new individuals with a genetic make-up resulting from the association and/or the reassortment of genetic material of different origins. In the most canonical form of sexual reproduction, the genome of an offspring, in other words the totality of its genetic material, derives from the union of (partial) copies of the genomes of two parents through fertilization. Here two distinct sexual processes are involved: the recombination of each parent’s genomes at meiosis during the production of gametes, and the combination of the genomes of the two gametes into the zygote’s genome at syngamy. Sexual reproduction involving two parents is called amphigony or biparental sexual reproduction. However, there are also forms of uniparental sexual reproduction (Chapter 6), in which the genome of a single parent is modified and reorganized during the process that leads to the offspring generation, as in parthenogenesis (reproduction through eggs that do not need to be fertilized) or in self-fertilization (reproduction through eggs that are fertilized by the sperm of the same individual). Sexual reproduction is found in the vast majority of multicellular eukaryotes and in most protists, but it does not necessarily rule out other forms of sex or other forms of reproduction.
With reference to the demographic and innovative aspects of reproduction mentioned in the previous section, the demarcation between sexual and asexual reproduction we have just established entails both the possibility of innovation without reproduction and without demographic growth (through sex alone), and the possibility of reproduction with or without sex, and with or without the production of genetic novelty.
Unfortunately, a discordant terminology is in use, based on different opinions on where to draw the line between sexual and asexual reproduction. In particular, what is contentious is whether some forms of uniparental reproduction, such as parthenogenesis, should be considered as sexual or asexual, along with the species or the individuals that practise them. In this book we have adopted a terminology that takes into account both the role of sex and the processes by which gametes are formed. Cases of uniparental reproduction involving sexual processes (as in self-fertilization), or deriving from processes typical of sexual reproduction (such as egg formation in parthenogenesis), are treated in the context of sexual reproduction, even when they have a clonal outcome. However, this terminological inconsistency in the literature is a minor point in comparison to the fact that there are forms of reproduction that challenge inclusion in any rigid classification. In any case, the reader is alerted: sexual and asexual may mean different things to different people.
Generations
We, the authors of this book, both belong to the ‘Boomer Generation’, people born in the two decades following World War II. On the Internet, this label is often used in contexts where teenagers and young adults (who evidently belong to another generation) tease attitudes typical of older adults. In the Western countries, the Boomer Generation was followed by Generation X, those born between the 1960s and the 1980s, then by the Millennials (Gen Y, born from early 1980s to mid-1990s), the Zoomers (Gen Z, born from mid-1990s to early 2010s,) and finally by the current generation Alpha. This is how sociologists and popular media subdivide generations: clusters of people born in a given period of time and, accordingly, expected to have experienced the same significant events of the society to which they belong. Most members of Gen Z are children of Gen X, not of the immediately preceding Gen Y. However, in biology (as well as in other sciences, like demography), the individuals born during the same breeding season or across the same interval of years are said to belong to the same cohort, while the term generation has a different meaning.
In biology, in particular in evolution and genetics studies, a generation is a set of individuals that come into being through a given number of reproductive events, either sexual or asexual, with reference to an individual ancestor or parent pair. Generations can thus be counted and numbered. For instance, many whale species travel in family pods that include three generations: a mother whale (parental generation), plus her daughter(s) (first offspring generation) and the young children of the latter (second offspring generation). All the descendants of an individual that reproduces several times during its life belong to the same generation. For instance, all the offspring of a female elephant that has been reproducing over multiple reproductive seasons form a single generation. Similarly, two seeds of sequoia produced by the same mother plant 2,000 years apart belong to the same generation (Figure 1.1).
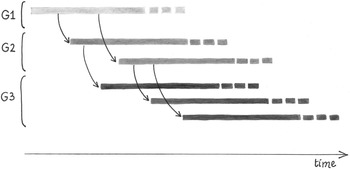
Figure 1.1. Schematic representation of the concept of generation. Horizontal thick lines are developing individuals belonging to three generations (G1–G3). Curved arrows are events of reproduction. Individuals produced at different times by different individuals, or even by the same individual, of the same (parental) generation belong to a single (offspring) generation. Note that individuals of a given generation may come into being before some individuals of a previous generation. For simplicity, a form of reproduction from a single parent is shown.
Belonging to a given generation is a relative characteristic, which depends on the choice of a reference individual ancestor, but another element of relativity is added in species with overlapping generations, where mating between individuals of different ages is possible. In his book L’ordine del tempo (The Order of Time), the Italian physicist Carlo Rovelli reconstructs king Leonidas’ family pedigree to illustrate the relativity of time in modern physics. This is a story of ‘generation relativity’.
Leonidas I, seventeenth king of Sparta, the hero of the Battle of Thermopylae during the second Persian War, married his half-brother’s daughter Gorgo and with her he had a son, Pleistarchus (Figure 1.2). Pleistarchus is at the same time son (first generation) and grandson (second generation) of two brothers. Gorgo belongs to the same generation as Leonidas (as parents), but the same can be said for her father Cleomenes, Leonidas’ brother (as children). With respect to Anaxandridas II, Leonidas’ and Cleomenes’ father, Pleistarchus belongs to the second generation through Leonidas and to the third generation through Cleomenes. Thus, generation assignment is not only relative to a given ancestor, but also relative to a specific ancestor–descendant path. The parent–offspring relationship establishes an order among some individuals belonging to a given set, but may not do so among all of them. Mathematicians use the term ‘partial order’ for the order provided by a relationship among the elements of a set that establishes an unambiguous precedence among some pairs of elements, but not among all pairs. Genealogy is an example of this kind of relationship.

This relativity in generation assignment shows up in all populations where individuals can reproduce across several breeding seasons, live long and become reproductively mature early, thus allowing overlap of generations, but the situation is even more embarrassing with some peculiar mating systems (Chapter 5). For instance, to which generation do the offspring born from the mating of a female with her own son (oedipal mating) belong? This is something that happens regularly in some organisms. Virgin females of the mite Histiostoma murchiei parasitize earthworm cocoons, in each of which they deposit 2–9 eggs that do not need to be fertilized. These will hatch within two or three weeks, producing only males. These males mature in about two days, mate with the mother, and die. The mother then lays about 500 fertilized eggs from which only females are born, which once developed will seek new earthworm cocoons to parasitize. These females are at the same time children and half-siblings (having the same mother) of their father, and children and grandchildren (being children of a child, their father) of the mother!
Living beings differ enormously in generation time, that is, in the average time interval between two consecutive generations. This interval varies from minutes for many bacteria (even as few as 12 minutes), to tens of years for large animals (20–30 years in our species) and plants (30–40 years in the beech, Fagus sylvatica).
Generation time has effects on the number of generations that can occur over a calendar year, or the number of years one generation may last. Small- and medium-size animals usually reach sexual maturity early and tend to have one generation per year (univoltine species) The same is true of many herbaceous plants (see below). This reproductive rhythm is particularly widespread among the species that inhabit regions with marked seasonality. However, there are numerous animals with particularly short generation times that have more than one generation per year (multivoltine species). The shortest generation time for an insect with amphigonic (i.e. biparental sexual) reproduction is a week for the mosquito Psorophora confinnis, while in case of parthenogenesis, generation time can be even shorter, less than five days, as in the aphid Rhopalosiphum padi.
In extreme cases of reproduction carried out in a juvenile stage (paedogenesis, Chapter 6), an individual can even begin to reproduce when it is still in the body of its mother. In some aphids, a parthenogenetic female carries in her body her developing daughters, and within these their own daughters (granddaughters of the former), like Russian dolls. Similar telescoped generations are found in some mites, in the stenolaematous bryozoans (marine invertebrates resembling colonial polyps), and in the colonial green alga Volvox.
In plants, depending on the length of the life cycle (which, however, as we will see, does not simply correspond to a single generation – see Chapter 2), we distinguish between annual plants, which complete the entire cycle within a year, like many herbs and grasses; biennial plants, which in the first year develop abundant foliage and bloom the following year, generally using the resources accumulated during the first year in bulbs, tubers or rhizomes; and perennials, which have longer life cycles. Annual and biennial plants are considered to have generation times of one and two years, respectively. In perennial plants the generation time depends on the age at which the plant reaches sexual maturity, which can vary from one year to several tens of years. Terrestrial plants able to complete several life cycles per year in natural conditions (corresponding to animal multivoltine species) are rare. Among these there are, in Europe, the alien populations of Galinsoga quadriradiata, a member of the sunflower family native to South America, which can complete 2–3 life cycles per year.
Reproduction and Individuality
Whatever way reproduction is defined, this implies accepting a suitable concept of a biological individual. Reproduction is the production of new entities, and these must correspond to some kind of individuals that can be counted. But what is an individual in biology? Rivers of ink have flowed in the attempts to answer this question, but the answer is that there is no unambiguous answer, for good biological reasons. We tend to conceive of individuals as being like ourselves, well-integrated entities, reasonably well defined in space and time, characterized by genetic homogeneity (all the cells in our body have the same genome) and genetic uniqueness (no one else has our genome, unless we have a twin), as well as by physiological unity and autonomy. However, that is simply not the case for many living beings.
Individuals lacking a unique genome are commonplace. Two amoebae that have just originated by fission from a parent amoeba share an almost identical genome, as do all the strawberry shoots derived from the same runner, and a pair of human identical twins. In all these cases, however, and particularly in the last one, we would be inclined to recognize these as distinct individuals. All forms of asexual reproduction and all forms of sexual reproduction with a clonal outcome undermine a definition of individuality based on genetic uniqueness. Asexual reproduction is very widespread among plants, so that botanists use two distinct terms to indicate two different kinds of ‘plant individuals’. A genet is a genetically unique entity, either a single physiologically independent entity, or a set of entities derived from a single individual by clonal multiplication (e.g. all the strawberry plantlets derived from the same runner). A ramet, instead, is an anatomically and physiologically bounded biological entity, with or without genetic uniqueness (e.g. any isolated strawberry plant, either derived from seed or from a runner) (Chapter 3).
As for the genetic uniformity of the individual, it is highly unlikely that the nuclei of the many cells of a multicellular organism have 100% identical genomes. When referring to the genetic identity among the members of a clone (such as the cells of a multicellular organism), it is implied that the mutations accumulated in the subsequent divisions starting from the founder cell are overlooked. The condition of an individual carrying different genomes that originated from the genome of a single founder cell is called genetic mosaicism. This common within-organism genetic heterogeneity, normally considered negligible, may become relevant in very old or very large organisms (i.e. those with many cells), in which the last common ancestor of two cells in the same individual’s body may be traced many cycles of cell division back. The most recent common ancestor of two reproductive cells of the same oak might have lived centuries ago, rather like the progenitor of a whole population of individuals in a species with an annual life cycle.
Within-organism genetic heterogeneity can also take the form of a genetic chimerism. A chimera (in biology, not in mythology) is a multicellular individual made of cell populations originating from more than one founder cell. Chimeric individuals can originate from the fusion of several spores in the red alga Gracilaria chilensis, from the fusion of larvae of the same species in freshwater sponges such as Spongilla, and a chimeric fungus can easily originate by fusion between hyphae of distinct individuals. But chimerism is also known among mammals. New World monkeys of the genus Callithrix, commonly known as marmosets, generally give birth to two (non-identical) twins. But these are not ‘normal’ twins. During pregnancy, connections between the two placentas are established so that cell exchanges occur between the two embryos. When the two little monkeys are born, each of them is a mixture of cells derived from the independent fertilizations of two distinct eggs. Thus, through this form of reproduction, two ‘genetic individuals’ and two ‘physiological individuals’ are obtained, but the two genetic individuals are distributed between the two physiological individuals. Furthermore, the two sperm that fertilized the two eggs can come from distinct fathers. This genetic promiscuity in parent–offspring relationships perhaps accounts for the peculiar and highly cooperative parental-care system in these monkeys. Possibly little known is the fact that a form of chimerism frequently affects our own species, together with other placental mammals. During pregnancy, fetal cells can migrate to the mother and vice versa. These cells can persist, multiply and even differentiate in the mother and the offspring for decades after birth (fetomaternal and maternofetal microchimerism).
A biological individual would be expected to be an undivided morphological and functional living unit, able to relate to the environment independently, including the ability to properly respond to environmental stimuli and the faculty to reproduce. However, individuals lacking autonomy and physiological unity are found among the members of highly integrated colonies, such as those of some marine invertebrates. Colonial hydrozoans known as the Portuguese man o’ war (Physalia) behave as an integrated unit to the point that the colony is often mistaken for an individual jellyfish. In each colony different types of individuals (zooids) coexist and cooperate, only some of which (gonozooids) are able to reproduce. In Volvox, a colonial green alga, only the special reproductive cells called gonidia, which are located within the sphere formed by flagellate somatic cells, can reproduce asexually to form new daughter colonies. Individuals lacking reproductive autonomy also include members of the sterile castes in some animal societies, like those of many species of bees, wasps, ants, termites and, unique among mammals, the naked mole-rat (Heterocephalus glaber).
Colonies and advanced societies pose a problem for the interpretation of the reproductive process, because the unit of reproduction can be identified at more than one level of biological organization – for instance, either in the single zooid of a colony or in the colony as a whole, which in the latter case can be seen as a ‘superorganism’.
Finally, autonomy becomes a myth when symbiosis is taken into account. Here the ‘boundaries’ of the individual definitely blur out. A large community of symbiotic microorganisms ‘inhabits’ the body of most multicellular organisms, which is necessary for their normal development and regular functioning. Our digestive system hosts a number of bacterial cells of the same order of magnitude as the cells (the ‘human cells’ in the strict sense) of our entire body (3.5 × 1013). In each person, these bacterial cells belong to a few hundred different ‘species’, out of the thousands which have been found in symbiosis with humans as a whole.
Beyond variation in attributes, individuals in the tree of life are not the same thing in another important respect: evolution. In their landmark book The Major Transitions in Evolution (1995), John Maynard Smith and Eörs Szathmáry analysed and discussed the recurrent events of emergence of new levels of organization in living systems. Multiple times, across a time span of more than two billion years, a certain number of entities originally able to survive and reproduce autonomously merged into a higher-level entity, causing a new hierarchical level of biological organization to appear. Among these events are the symbiotic union of primitive prokaryotic cells in the first eukaryotic cell, the evolution of multicellularity in several lineages of eukaryotic unicellular organisms, the evolution of colonial organisms and animal societies. Thus, a bee colony is an individual made of thousands of organisms, each made of billions of eukaryotic cells, each originally derived by the fusion of two prokaryotic cells. These ‘major evolutionary transitions’ are actually ‘evolutionary transitions in individuality’. At each one of these events, a new kind of individual has evolved through cooperation and integration of pre-existing individuals. However, these new individuals have not replaced the older ones. Rather, they have added a new level of individuality in the tree of life, increasing its diversity at this fundamental level.
Summing up, there are different kinds of individual out there. This may seem a complication for our schemes and models, but this is life, always intolerant of our simplistic definitions. However, we can cope with that. Keeping a flexible attitude towards the concept of an individual will be important in the following chapters, where we will see reproduction at work in different kinds of organism. The kind of individual and the mode of reproduction will define the boundaries between reproduction and other biological processes occurring in the same organism, like growth or development, but these boundaries are not always neat. We will highlight these ‘difficult boundaries’ along the way, to return with some final considerations towards the end of the book.
Reproduction and Senescence
Reproduction allows the persistence of a species despite the continual loss of its individual members. Mortality rate per unit of time in a population is never zero. Individual living beings do not last forever; they are mortal for two main reasons. First, an individual may die by an accident related to its interactions with predators or parasites, or to environmental physical factors beyond its tolerance threshold. Second, even in environmental conditions ideal for its survival, with no limit to the availability of resources for its maintenance, the individual cannot save itself from ‘certain death’. This is the inescapable result of the developmental process called senescence (or ageing), manifested with an increase in the probability of death with age. Senescence is a cumulative process, occurring at different levels of body organization (from molecules to cells, tissues and organs), which progressively or abruptly, depending on the species, corrupts metabolism and body structures, producing a deterioration of the qualities of the organism that eventually leads to its death. In some cases, as in many annual plants, the squids and the Pacific salmon, senescence is triggered quickly the first and only time the organism reproduces sexually.
Senescence is the antagonist of immortality. In biology, to be immortal does not mean that an organism cannot die, a quality reserved for certain mythological figures and comic-book superheroes. All living things may die from trauma, disease, or simply being eaten by another living being. In biology, immortality, with reference to either a cell or an individual, is rather a potentiality. It is the absence or the arrest of senescence. Whereas living beings may all die, senescence does not affect all organisms in the same way. An individual or cell that does not age, or ceases to age at some point in its existence, is said to be biologically immortal.
Most prokaryotes and many protists do not seem to experience senescence. Also, there is no certain evidence of senescence in some plants that live for a very long time, over 4,000 years, including some conifers (Pinus longaeva and Sequoiadendron giganteum), some sponges and sea anemones, hydras, the queens of different species of social insects (bees, ants, wasps and termites), some tube-dwelling polychaetes (Lamellibrachia) and certain bivalves (Arctica). The same may apply to the black coral Leiopathes, for which a maximum age of 4,265 years has been estimated. All these organisms are considered potentially immortal.
For all the living beings that age, it is not enough for reproduction to generate new individuals, in addition to those already present, or to replace those that have died. Reproduction must also ensure that newborns are actually ‘young’ – in other words, that they have, so to speak, ‘turned back the clock of senescence’ so that the population ‘rejuvenates’ through reproduction. To generate young individuals from old ones is an imperative for the continuity of life. This is accomplished in many different ways. Sexual reproduction has this ability to rejuvenate. Through the processes that lead to the formation of gametes in multicellular organisms, for instance, the senescence timer is effectively reset to zero. The life expectancy of a fertilized egg is definitely higher than the life expectancy of the two parents from which the gametes were produced. Within certain limits, varying from species to species, it is also independent of the parents’ age.
The capacity of rejuvenation seems to be an attribute also of sex in the broad sense. Ciliates reproduce only asexually, in many species by simply dividing the cell into two (binary fission, Chapter 3), but commonly practise conjugation (see above), a form of sex where two individuals (conjugants) come into contact, mix their genetic material, then separate again (as ex-conjugants), genetically different from those that conjugated, but identical to each other.
In most ciliates, the clone that originates from an ex-conjugant after separating from its partner shows a form of clonal senescence, consisting of a limit to the number of cell divisions it can undergo. This number varies from species to species, but also between strains of the same species. In Tetrahymena this limit varies between 40 and 1,500 divisions. Moreover, the clone goes through different ‘maturation stages’ comparable to what in a multicellular organism we might describe as developmental phases. During an initial period of ‘sexual immaturity’ of the clone (measured by the number of divisions since the last conjugation), individuals can only multiply asexually, but are not able to conjugate. Then follows a period of ‘sexual maturity’ during which they will be able again to conjugate. Ex-conjugants will emerge from this event genetically modified, but also in some way rejuvenated, with an expected number of future cell divisions equal to the maximum possible for the species or strain. Individuals that do not conjugate, instead, may continue to multiply, but will enter a phase of gradual senescence: the rate of cell divisions will gradually slow down, eventually leading to the extinction of the clone. If they conjugate during this phase of senescence, the ex-conjugants will have an expectancy of clonal propagation below the maximum value for the species.
In some cases at least, asexual reproduction generates rejuvenated offspring as well. Examples are provided by many plants with vegetative reproduction. A branch broken away from an old willow has a good chance of taking root and developing into a new young individual with a life expectancy that does not depend on the age of the parent. There are clones of quaking aspen (Populus tremuloides) estimated to be more than 80,000 years old (see Concluding Remarks), of the creosote bush (Larrea tridentata) almost 12,000 years old, and of bracken (Pteridium aquilinum) almost 1,500 years old.
In unicellular organisms, prokaryote and eukaryote alike, the unequal partition of damaged cell constituents among the daughter cells, even if cell division is apparently symmetrical, can rejuvenate the clone through the propagation of damage-free cells. For instance, individual cells of the yeast Saccharomyces cerevisiae divide asymmetrically, so that the larger cell is called the mother cell, and the smaller the daughter cell. The mother cell is subject to a form of senescence by accumulating damaged components, so that there is a limit to the number of daughter cells (about 50) it is able to produce before finally ceasing to divide. Until a certain ‘age’ of the mother cell (number of divisions undergone till then), the daughter cells will not receive any damaged components from the mother, and the replicative potential (number of possible divisions) of a daughter cell, which will soon act in turn as a mother cell, will be the maximum for the species. However, as the age of the mother cell increases, the daughter cells will receive increasing quantities of damaged components, thus detaching from the mother cell with a replicative potential already reduced compared to the species’ maximum (Figure 1.3).

Figure 1.3. Asymmetric clonal senescence in the yeast Saccharomyces cerevisiae. Along the series of budding events for the same individual cell, the mother cell progressively accumulates senescence factors (crosses). Daughter cells progressively receive larger quantities of these factors. Daughter cells that are generated early retain full replication potential, while daughter cells generated later will emerge from the division of the mother cell with a reduced replication potential.
Reproduction and Us
We share many aspects of reproduction with many other organisms. We reproduce sexually, we have chromosomal sex determination (Chapter 7), parental care ( Chapter 8) and a diplontic life cycle (Chapter 2). We also practise some forms of courtship, although, uniquely in the living world, this can now happen through the screen of a smartphone. However, in some way (or in many ways, depending on the viewpoint) we are special, or at least we think we are. This impression is not totally unjustified. Beyond any form of human chauvinism (e.g. ‘we write books about other species, but no other species writes books about us’), there are two ways in which we are objectively special: one is generic, the other … special. We are special in the generic sense that every species is special. In this book, we will explore what is common among the ways organisms reproduce, but in the end every species is a separate case, and generalizations can miss some unique feature of reproduction in any given species. The other, special sense in which our species is special is that this is our own species. When we deal with human biology, we are dealing with ourselves, and the way we can study, investigate and understand what we do has no comparison with what we can do on other species. For instance, physicians, psychologists and sociologists use in their studies the answers given by a sample of subjects to a set of questions they are asked. We also assume that important aspects of perception and consciousness are shared among all of us. These aspects are an intimate component of any biological phenomenon that regards ourselves, but their study and understanding cannot be extended to any other living beings.
The relatively simple question of how many sexual conditions there are in a given species can have a straight answer for squirrels (two: male and female), for garden snails (one: hermaphrodite) or for certain populations of the wheel cactus (Opuntia robusta) (three: male, female and hermaphrodite). The sexual condition is defined on the basis of the kind of gametes an individual organism can produce (sperm, eggs or both, Chapter 4), and in principle there is no ambiguity in assigning a given individual to one of the three categories, male, female or hermaphrodite. However, as you can imagine, the two simple questions of how many sexual conditions can be recognized in humans and, for a given individual, to which sex does s/he belong, can touch on very delicate issues that are the subject of psychology, anthropology and sociology. In other species, an individual with an anomalous combination of male and female features, or with sex-specific features in an intermediate form, is labelled a gynandromorph or an intersex (Chapter 7). We can study the genetics, morphology, physiology and behaviour of these individuals. For instance, a gynandromorph zebra finch, male on the right side, female on the left side, sang a fully masculine song and courted and copulated with a female (in birds, the right brain hemisphere controls social behaviour). However, we cannot ask the bird how it felt, or how it identified itself.
In most separate-sex species, the sex condition of a given individual can be complicated by the fact that both the establishing of the sex of an individual (sex determination) and the emergence of its sex characters (sex differentiation) are developmental processes, often very complex (Chapter 7). Thus, it is possible that the ‘genetic sex’ of an individual (e.g. its sex-chromosome complement) is at odds with the titre of sex hormones circulating in the blood (‘hormonal sex’), or with the structure of the gonad (‘gonadal sex’). Moreover, some sexual traits can be intermediate between male and female, or the individual may show a mosaic of male and female traits, either morphological or behavioural. This is already complex enough; however, in our own species additional questions emerge, involving psychological and sociological factors.
To treat these questions in humans, a common terminological contrivance is to distinguish between sex and gender, where sex includes ‘biological’ attributes like genitalia, sex-related chromosomes and hormones, while gender includes the individual’s psychological and social attributes associated with maleness and femaleness. Gender identity is about how a person identifies, but does not come in binary form, since gender embraces a broad spectrum of possibilities, and a person may identify at any point within this spectrum or even outside of it. It is also recognized that gender identity can change multiple times during a life. Transgender, non-binary, gender-fluid, gender-neutral are only examples of the variety of ways in which a person may define their own situation or viewpoint. Gender orientation describes an individual’s physical and/or emotional attraction to other individuals, where heterosexuality, homosexuality and bisexuality are the most common options. Gender is also a social affair. Belonging to a given gender category entails different expectations or behavioural models of, say, masculinity and femininity in different societies, and some people do not identify with some, or any, of the features associated with their gender. Although in most societies there is a basic division between gender attributes assigned to males and females, respectively, some societies have one or more additional gender categories, as some anthropologists and sociologists have described. However, practical as it may seem, the distinction between sex and gender is not universally accepted. Some scholars see a difficulty separating the variables that should discriminate between the physical and the psychological spheres, and tend rather to stress the intimate connection between the two. Gender identity and orientation presumably emerge from a multitude of bodily aspects via (poorly understood) interactions with environment and experience. In any case, the term ‘gender’ should not be used in non-human species.
Whatever gender may mean, we will not touch on ‘gender questions’ in this book. This is not because they are not part of our biology, but because they involve mental faculties, such as consciousness, self-perception, intention and will, which, although possibly not exclusive to the human species, are not accessible to investigation in the same way in other species, or through the same analytical tools applicable to our species. It is a question too specific to our own species, and this book is about what is common among the ways all organisms reproduce, with no special focus on human reproduction and sexuality. However, we need to clarify two things, to stress the difference between discussing sex and related matters in our species and in all the others.
First, as a sort of disclaimer, the contents of this book should not be taken as a biological foundation supporting any particular moral, social or political arguments about gender questions, human rights or related medical practices. Observing and studying nature is always informative, but we should be careful not to take nature as a model for humans. Thinking that nature, which in this case equates to the biology of other organisms, can provide a guide for our moral judgement or for taking decisions about our social ordering is a logical error so common that it has a name – the naturalistic fallacy or fallacy of the recourse to nature. This may prompt questions of whether homosexuality, polygamy or incest (but also xenophobia or aggression) are natural, or if they are not instead the (degenerate) product of our culture. In doing so, a double mistake is made, by separating nature and culture, forgetting that culture is an intimate part of human nature, like flight to a sparrow, and by picking up and giving credit to the natural cases that support our (pre)moral judgements, while ignoring those that contradict them. Just to mention the three examples above, homosexuality has been described in some species (e.g. the Chilean flamingo) but not in others; there are species where mating promiscuity is the norm (e.g. the chimpanzee), while in others a pair-bond lasts forever (e.g. swans); there are cases where mother–son mating (e.g. some roundworms) or brother–sister mating (e.g. some mites) is the norm, but this does not occur in all species. But there are also siblings that eat each other (fratricide, many raptors), offspring that devour their mother (matricide, some spiders), females that kill and eat their partner (mariticide, mantises), males in a group that harass a female to mate with her (group rape, some whales), and this list could go on. What should we take from this for ourselves? Nothing, probably, other than an appreciation of some unexpected facets of the diversity of life’s phenomena.
Second, the language we adopt follows the scientific use of the terms involved. When we illustrate in later chapters the possibility of ‘sex without reproduction’ and ‘reproduction without sex’, one might mistakenly understand that we are talking about contraception and in-vitro fertilization, respectively. But what we actually mean here is that there are sexual processes not associated with reproduction, and modes of reproduction that do not involve sexual processes. When we say ‘sexuality’ we are referring to sex process, not to the looser behavioural concept of the ‘sexual sphere’ in our species. And, again, the terms ‘abnormal’ and ‘normal’ do not have any connotation as pathological versus non-pathological (as in clinical medicine), and do not take any aesthetic or ethical value (as less-than-ideal versus ideal). The term ‘normal’ generally refers to the most commonly encountered variant among a set of possibilities, corresponding to the modal type/condition in statistics. This is the way a sentence like ‘in separate-sex species, gynandromorphs are abnormal individuals with a mix of male and female traits’ should be understood.
That said about ourselves, let’s zoom out again, to regain a wider whole-tree-of-life perspective.
The Theoretical Minimum
The great Russian physicist Lev Davidovich Landau (1908–1968) called ‘theoretical minimum’ everything a student needed to know to work under his tutorship. Landau was a very demanding professor: his theoretical minimum meant just about everything he knew, which of course no one else could possibly know. Thankfully, years later Leonard Susskind and George Hrabovsky used the same term with a different, more accessible meaning. The theoretical minimum that gives their best-selling introduction to physics the title is just what you need to know in order to proceed to the next level. In the same vein, we provide here the minimum of biological knowledge to understand what we will talk about in the following pages. If you have already studied basic biology, you can skip this section.
Living organisms, in a sense that excludes viruses, are made of cells: just one, as in bacteria, ciliates and amoebae, which are thus described as unicellular; or many, up to thousands of trillions in a big whale, in organisms which are described as multicellular. In multicellular organisms, many different cell types occur in the body of a single individual, to form specific tissues and organs. For example, no less than 200 types are recognized in the human body.
In terms of structure, cells are of two fundamental types. On the one hand, there is the prokaryotic cell, found in bacteria and bacteria-like organisms, which is small (size order 1 μm), with no internal organelles and DNA not separated from the other contents of the cell. On the other hand, there is the eukaryotic cell, typical of both unicellular and multicellular eukaryotes; it is larger (size order 10 μm), and with complex content, which includes a nucleus (a closed envelope that encloses most of the genetic material), membranes and organelles.
The totality of the genetic material of a cell or of an organism, in the form of DNA molecules, is its genome. In eukaryotic cells, the DNA molecules in the nucleus, or chromosomes, carry by far the most conspicuous part of the genome, but not its totality. In addition to the nuclear genome there are smaller genomes associated with two kinds of organelles: the mitochondria, the site of energetic processes known as cellular respiration, which are present in virtually all eukaryotes, and the plastids, characteristic of plants, including algae, which contain chlorophyll or other pigments involved in photosynthesis. These organelles are the remnants of bacteria, originally free-living, that have been incorporated a long time ago by a larger cell – within which they survive in a relation of symbiosis, preserving a degree of autonomy in function, replication and transmission across generations, as shown by the persistence of some part of the original bacterial chromosome.
Chromosomes are described as carriers of genes, segments of DNA involved in the development of traits. Alternative forms of the same gene (or, equivalently, at the same genetic locus) are called alleles. The whole set of gene alleles in a cell (or in the organism) is its genotype. The latter term is contrasted to phenotype, the whole set of its observable characteristics, morphological especially.
An important feature of the organization of the genetic material is the number of complete sets of chromosomes in the nucleus, which are generally one or two. Homologous chromosomes in the different sets have the same genes, although each gene can be present with different alleles. In a human cell, for examples, there are two sets of 23 chromosomes, and thus we are diploid; but the unicellular alga Chlamydomonas is haploid (one set of chromosomes) and the parthenogenetic Indo-Pacific gecko (Hemidactylus garnotii) is triploid (three sets). A condition with more than two sets is called polyploidy. A cell, or an organism, that presents the same alleles at a given locus of the homologous chromosomes is said to be homozygous at that locus; alternatively, it is said to be heterozygous.
In prokaryotes, cell division occurs simply by cell fission after the DNA has been duplicated, but in eukaryotes the division of the nucleus, followed by the splitting of the cell, can follow two fundamentally different routes (Figure 1.4). With mitosis, chromosomes are first replicated and then equally distributed into two new nuclei, followed by the division of the cell. The two resulting cells are genetically identical and the total number of sets of chromosomes is maintained. In meiosis, after chromosome replication, two rounds of chromosome separation follow, which can result in four nuclei, and generally in as many cells, each with half the original number of chromosome sets. In the sexual reproduction of diploid organisms, like us, meiosis gives rise to the haploid egg and sperm from diploid germ cells. Importantly, during meiosis, there is reshuffling of genetic material (recombination) so that the chromosomes in the gametes are a mix of the genetic information contained in the two sets of chromosomes, which as a rule were independently inherited by the two parents. Both mitosis and meiosis feature prominently in the life cycle of most eukaryotes (we will have a lot to say about that in this book), but there is another fundamental process that allows the re-establishment of the original chromosome number after meiosis. This is karyogamy, the fusion of two nuclei and their chromosomes, as typically follows the fusion of gametes, or syngamy. The resulting cell, called a zygote, can be the founder cell for the development (through mitosis) of a multicellular organism. As we will see, in most animals, meiosis is the process by which eggs and sperm cells are produced, but in other kinds of organisms the relationships between meiosis and karyogamy are not the same.

In the next chapter we will take a look at the context in which reproduction occurs, the organism’s life cycle.




