


Introduction
We present a temporally calibrated, shallow marine δ18O chemostratigraphic curve for Bentiaba, southern Angola (Fig. 1), relating stable oxygen isotopes of this portion of the southern hemisphere to values from the Walvis Ridge offshore Angola and elsewhere around the globe. The presented δ18O values are used to estimate the paleotemperature regime in which mosasaurs and other marine amniotes along this portion of southwest Africa lived throughout much of the Late Cretaceous.

Fig. 1. Location map of Bentiaba, Angola. Inset shows location of Angola within Africa, offshore basins labelled in capital letters. Note location of DSDP Hole 530A, which is discussed in the text. Modified from Strganac et al. (Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a).
Mosasaurs have a fossil record extending from the Cenomanian, approximately 98 Ma, to their extinction at the end of the Cretaceous at 66 Ma (Jacobs et al., Reference Jacobs, Ferguson, Polcyn and Rennison2005a,b; Polcyn et al., Reference Polcyn, Tchernov and Jacobs1999, Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014). Since 2005 our team, Projecto PaleoAngola, has conducted field expeditions in Cretaceous and younger rocks of coastal Angola, greatly improving the fossil record of vertebrates from the eastern South Atlantic (Jacobs et al., Reference Jacobs, Mateus, Polcyn, Schulp, Antunes, Morais and Tavares2006). The oldest mosasaurs known from the South Atlantic Ocean, Angolasaurus bocagei and Tylosaurus iembeensis, are found at Iembe, north of the capital Luanda, in Late Turonian nearshore deposits that also produced the sauropod dinosaur Angolatitan adamastor (Mateus et al., Reference Mateus, Jacobs, Schulp, Polcyn, Tavares, Neto, Morais and Antunes2011). Marine amniote remains known from Iembe also include halisaurine vertebrae and a marine turtle, Angolachely mbaxi (Mateus et al., Reference Mateus, Jacobs, Polcyn, Schulp, Vineyard, Antunes and Neto2009, Reference Mateus, Polcyn, Jacobs, Araújo, Schulp, Marinheiro, Oereira and Vineyard2012; Polcyn et al., Reference Polcyn, Lindgren, Bardet, Cornelissen, Verding and Schulp2012).
The greatest concentration of mosasaurs in Angola comes from Upper Campanian–Maastrichtian sediments at Bentiaba (Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014b), but there are significant exposures below the fossil-bearing horizons, which range in age from Cenomanian to Santonian. The chronological context for Bentiaba is provided by the magnetostratigraphy and a δ13C chemostratigraphic framework anchored by an 84.5 Ma 40Ar/39Ar whole-rock radiometric date on an intercalated basalt of the Ombe Formation, which is consistent with ammonite biostratigraphy (Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a). The Bentiaba δ18O curve presented here was constructed as a complement to the δ13C record presented by Strganac et al. (Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a).
The mosasaur fauna of the Campanian Baba Formation is fragmentary and includes both russellosaurians and mosasaurines, while that of the overlying Maastrichtian Mocuio Formation is predominately mosasaurine (Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014b). Within the Mocuio Formation, the mid-Maastrichtian (approximately 71.5 Ma) Bench 19 fauna is particularly rich (Schulp et al., Reference Schulp, Polcyn, Mateus, Jacobs, Morais and Tavares2006, Reference Schulp, Polcyn, Mateus, Jacobs, Morais and Everhart2008, Reference Schulp, Polcyn, Mateus and Jacobs2013; Polcyn et al., Reference Polcyn, Jacobs, Schulp and Mateus2010, Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014; Mateus et al., Reference Mateus, Polcyn, Jacobs, Araújo, Schulp, Marinheiro, Oereira and Vineyard2012), and it is the subject of a paleoecological analysis (Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014b). The paleogeographic context of Angolan vertebrates was initially examined in Jacobs et al. (Reference Jacobs, Mateus, Polcyn, Schulp, Scotese, Goswami, Ferguson, Robbins, Vineyard and Neto2009, Reference Jacobs, Strganac and Scotese2011) and Polcyn et al. (Reference Polcyn, Jacobs, Schulp and Mateus2010), and their paleoecology was examined initially in Robbins et al. (Reference Robbins, Ferguson, Polcyn, Jacobs and Everhart2008) and Polcyn et al. (Reference Polcyn, Jacobs, Schulp and Mateus2010, Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014).
Little paleotemperature research relevant to this study has been conducted in Africa. An analysis of paleosols in the Samba drill core from the Congo Basin (Cahen et al., Reference Cahen, Ferrand, Haarsma, Lepersonne and Verbeek1959), representing the interior of Gondwana prior to the opening of the South Atlantic, indicates Jurassic average annual soil temperatures between 25 and 40±3°C, with an average of 33°C, σ = 7.6°C (Myers et al., Reference Myers, Tabor and Jacobs2011), accompanied by low biological productivity (Myers et al., Reference Myers, Tabor, Jacobs and Mateus2012). Myers et al. (Reference Myers, Tabor and Jacobs2011) also presented a Lower Cretaceous soil temperature estimate of 23±3°C.
Temperature tolerances for mosasaurs of Cenomanian age, near the time of origin for the group, were considered by Jacobs et al. (Reference Jacobs, Polcyn, Taylor and Ferguson2005b). Kolodny & Luz (Reference Kolodny, Luz, Taylor, O’Neil and Kaplan1991) reported the δ18O value of the Cenomanian shark Cretolamna appendiculata from Angola, included by Pucéat et al. (Reference Pucéat, Lécuyer, Donnadieu, Naveau, Cappetta, Ramstein, Huber and Kriwet2007) in their study of Cretaceous latitudinal temperature gradients. The current study provides temperature estimates of shallow marine environments, below wave base, for much of Late Cretaceous time as the South Atlantic was growing and was inhabited by diverse assemblages of marine amniotes.
Cretaceous δ18O records have been used to elucidate changes in ocean circulation as well as temperature (Cramer et al., Reference Cramer, Toggweiler, Wright, Katz and Miller2009; Friedrich et al., Reference Friedrich, Herrle, Wilson, Cooper, Erbacher and Hemleben2009, Reference Friedrich, Norris and Erbacher2012). Most paleotemperature records utilise benthic foraminifera, whose δ18O values reflect offshore deep marine environments with little to no influence by meteoric input or fluctuations in salinity (Grossman, Reference Grossman, Gradstein, Ogg, Schmitz and Ogg2012). These methods can be extended to shallow shelf setting unaffected by high freshwater influx (Sessa et al., Reference Sessa, Ivany, Schlossnagle, Samson and Schellenberg2012), which we assume to be the case at Bentiaba (Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014b). The δ18O curve from Bentiaba provides a shallow marine temperature curve that can be compared to the long-term benthic records to track broader temperature trends, to obtain indications of larger circulation patterns, and to provide a paleotemperature context for evolution in the marine realm.
The Late Cretaceous marks the transition from global high temperatures during the Late Cenomanian-Turonian climatic maximum and progressing through the cooler temperatures that characterise much of the Cenozoic. Cretaceous temperature trends coincide with significant paleoceanographic events, including Oceanic Anoxic Event 2 at the Cenomanian-Turonian Boundary, which is a widespread period of rapid burial of organic matter related to marine productivity and enhanced ocean stratification (Arthur et al., Reference Arthur, Dean and Pratt1988; Erbacher et al., Reference Erbacher, Huber, Norris and Markey2001, Forster et al., Reference Forster, Kuypers, Turgeon, Brumsack, Petrizzo and Sinninghe Damsté2008; Friedrich et al., Reference Friedrich, Norris and Erbacher2012; Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a). Connections between major ocean basins increased after the Turonian as a result of tectonic drift of continents, allowing cool waters derived from high latitudes to circulate equatorially, contributing to declining global temperature (Hay et al., Reference Hay, DeConto, Wold, Wilson, Voigt, Schultz, Wold, Dullo, Ronov, Balukhovsky, Söding, Barrera and Johnson1999; Miller et al., Reference Miller, Barrera, Olsson, Sugarman and Savin1999; Cramer et al., Reference Cramer, Toggweiler, Wright, Katz and Miller2009; Robinson & Vance, Reference Robinson and Vance2012). The δ18O stratigraphic curve presented here is a proxy for shallow marine temperature change at Bentiaba during the Late Cretaceous development of the South Atlantic.
The mosasaur-bearing portion of the Bentiaba section lies above the Ombe basalt in the Baba and Mocuio formations (Fig. 2). Carbonate cement creates benches in the Mocuio Formation, which are numbered in the section. Vertebrates occur throughout the Baba and Mocuio formations. The Campanian Baba Formation preserves a fragmentary but mixed russellosaurine-mosasaurine fauna extending into the lower part of the Mocuio Formation. In the Lower Maastrichtian part of the Mocuio Formation, the Bench 19 Fauna is dominated by mosasaurines. The highest levels of the Mocuio fauna, which we interpret as Upper Maastrichtian (Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a), preserve the globidensine Carinodens and a large, derived Prognathodon (Mateus et al., Reference Mateus, Polcyn, Jacobs, Araújo, Schulp, Marinheiro, Oereira and Vineyard2012; Schulp et al., Reference Schulp, Polcyn, Mateus and Jacobs2013; Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014b).

Fig. 2. Section at Bentiaba with δ13C and inoceramid δ18O stratigraphic curves. Red bones and ‘19’ indicate marine amniote-bearing horizons and Bench 19. Note that the δ13C curve is composite of inoceramid and other bivalve data in Strganac et al. (Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a).
Methods and materials
Stratigraphic δ18O sampling
The inoceramid bivalves in this study represent a subset of data used in a δ13C chemostratigraphic curve at Bentiaba (Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a). Marine bivalves throughout the section were sampled for stable isotope chemostratigraphy. Strganac et al. (Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a) tested inoceramid shells for diagenetic alteration by visual examination with binocular microscope and by scanning electron microscopy (SEM). Additional tests for alteration were performed by electron dispersive spectroscopy (EDS) and X-ray diffraction. Only samples of inoceramids with intact calcite prisms were used to estimate paleotemperatures in this study. Samples altered to dolomite were rejected, and stable oxygen isotope values from non-inoceramid bivalves were not used in this study, as these samples were not examined for diagenetic alteration in detail.
Twenty-four inoceramid shells were powdered with a carbide drill near the hinge where the shell is thickest to develop δ13C and δ18O chemostratigraphic curves. These powder samples were vacuum-sealed in vessels and reacted with 100% orthophosphoric acid. Calcite reactions were performed in a 25°C bath for at least 4 hours and dolomite was reacted at 50°C for at least 24 hours. The resulting carbon dioxide gas was cryogenically purified and analysed in a Finnigan MAT 252 mass spectrometer with a precision of 0.1‰.
The δ18O values were used to produce an oxygen isotope chemostratigraphic curve for Bentiaba that was then compared to δ18O compilations from the South Atlantic at Walvis Ridge and from the Pacific Ocean (Friedrich et al., Reference Friedrich, Norris and Erbacher2012). Paleotemperatures (T) were estimated using the calcite-temperature equation of Kim & O’Neil (Reference Kim and O’Neil1997):
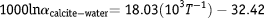 $${\rm{1000ln}}\alpha _{{\rm{calcite - water}}} {\rm{ = 18}}{\rm{.03(10}}^{\rm{3}} T^{{\rm{ - 1}}} {\rm{)}}-{\rm{32}}{\rm{.42}}$$
$${\rm{1000ln}}\alpha _{{\rm{calcite - water}}} {\rm{ = 18}}{\rm{.03(10}}^{\rm{3}} T^{{\rm{ - 1}}} {\rm{)}}-{\rm{32}}{\rm{.42}}$$
where α calcite-water represents the calcite–water fractionation factor.
Results
The results of the stable oxygen isotopic analysis for 14 calcite shells and 10 additional samples that were altered to dolomite are shown in Table 1. The δ18O values of calcite samples range between –3.2‰ and –0.8‰, with an average value of –1.6‰. These values and those for shells altered to dolomite are plotted in Fig. 2 against stratigraphy and the δ13C chemostratigraphic and paleomagnetic data from Strganac et al. (Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a). The resulting pattern in stable oxygen isotopes of calcite shows a general trend of increasing δ18O values up section. A positive ∼2‰ excursion in the uppermost Salinas Formation to Lower Baba Formation results from values of –3.2‰, –2.1‰ and –1.2‰ derived from shells recovered at 57, 67 and 69.5 m, respectively. Between 71 and 81 m, δ18O values remain relatively constant, ranging between –1.4 and –1.9‰. The maximum values of –0.8‰ and –0.9‰ are from shells collected at 108 m and 110.5 m. A positive 0.5‰ shift is observed in inoceramids recovered at the base of the Mocuio Formation, at 116 m, and remain between –1.2‰ and –1.8‰ to 128 m.
Table 1. Stable isotope values of inoceramid shells for the Bentiaba δ18O stratigraphy. Paleotemperature calculated using calcite–temperature equation of Kim & O’Neil (Reference Kim and O’Neil1997).

Discussion and conclusions
The pattern of δ18O from inoceramids at Bentiaba is similar to those known from benthic foraminifera from the Pacific Ocean, albeit with a negative ∼1‰ offset (Fig. 3). This suggests that the oxygen isotope stratigraphy at Bentiaba tracks declining global temperatures beginning in the Late Turonian, concurrent with a deep ocean connection in the equatorial Atlantic. A δ18O value of –3.2‰ derived from inoceramids at Bentiaba coincides with depleted values during the Late Turonian from benthic foraminifera in the Pacific and Inoceramus at Walvis Ridge. Values decrease ∼2‰ by the Late Campanian, similar to the decrease observed at Walvis Ridge (Barron et al., Reference Barron, Saltzman and Price1984). In the Late Campanian, fluctuations of less than 1‰ in δ18O at Bentiaba may reflect smaller variability in temperature or effects of salinity superimposed on a warming interval observed globally. Similar fluctuations in δ18O values are also observed near the same time interval at the Walvis Ridge (Barron et al., Reference Barron, Saltzman and Price1984).

Fig. 3. Comparison of Bentiaba δ18O stratigraphy to Inoceramus (Barron et al., Reference Barron, Saltzman and Price1984) and benthic foraminifera (Li & Keller, Reference Li and Keller1998; Friedrich et al., Reference Friedrich, Norris and Erbacher2012) from Walvis Ridge and the Pacific Ocean compilation (Friedrich et al., Reference Friedrich, Norris and Erbacher2012; their supplemental). Figure is as follows left to right: Bentiaba section with stable oxygen- and magnetostratigraphy, Bentiaba δ18O curve calibrated with δ13C (horizontal shaded areas) and paleomagnetic (dashed lines) correlations from Strganac et al. (Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a), Walvis Ridge and Pacific δ18O curves.
Bentiaba δ18O values indicate a similar pattern to global values from the Turonian through the Late Campanian (Fig. 3) during a time when the shallow marine environment at Bentiaba became increasingly confined, possibly due to falling sea levels during the Campanian, indicated by dolomitisation of inoceramids at two stratigraphic levels in the Bentiaba section during the Late Campanian and Early Maastrichtian (Warren, Reference Warren2000; Friedrich et al., Reference Friedrich, Herrle, Wilson, Cooper, Erbacher and Hemleben2009; Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a). While these conditions may seem to support higher evaporation rates and high salinities, the relatively depleted values at Bentiaba, offset by negative ∼1‰ from deep ocean values, does not support this. Barron et al. (Reference Barron, Saltzman and Price1984) predicted that the enrichment in δ18O at Walvis Ridge in the Campanian was the result of dense saline waters originating from shallow seas from Africa, which our results also do not support. Depleted δ18O values may result from meteoric input, but Bentiaba was within the arid latitudes (∼24°S) and the sedimentary textures, mineralogy and sources suggest low meteoric input (Strganac et al., Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014b).
At Bentiaba, a Late Turonian paleotemperature estimate from calcite is ∼25°C. In the earliest Campanian the estimated paleotemperatures decrease to 15–18°C, similar to that estimated for the Australian Pacific (Friedrich et al., Reference Friedrich, Norris and Erbacher2012). By the Late Campanian–earliest Maastrichtian, Bentiaba paleotemperatures remain approximately the same, 16–19°C, whereas temperatures in the Pacific and Walvis Ridge have decreased to ∼10°C. Assuming a constant ocean δ18O value during the Cretaceous, the change in δ18O values from –3.2 to –1.2‰ observed at Bentiaba corresponds to a ∼10° temperature decrease from the Late Turonian to Early Maastrichtian, similar to temperature changes inferred from oxygen isotopes derived from deep sea benthic foraminifera in the Pacific Ocean and Southern Ocean (Clarke & Jenkyns, Reference Clarke and Jenkyns1999; Cramer et al., Reference Cramer, Toggweiler, Wright, Katz and Miller2009; Friedrich et al., Reference Friedrich, Herrle, Wilson, Cooper, Erbacher and Hemleben2009, Reference Friedrich, Norris and Erbacher2012).
Mosasaurs are known from Angola from the Late Turonian at Iembe to the Maastrichtian at Bentiaba (Fig. 1), during a trend of declining sea temperature implied by the Bentiaba stable oxygen isotope curve. During the Turonian, temperatures at Bentiaba ranged from ∼22.4 to 25.3°C (Table 1). The temperature calculated from δ18O values at the lowest amniote fossils at Bentiaba (Baba Formation, Chron C33n, approximately 75 Ma) is 16.7°C. The temperature estimate for Chron C32n2n (approximately 73 Ma), within the Baba Formation, which contains indeterminate russellosaurian and mosasaurine mosasaur fossils, is 19.0°C. The estimate for the mosasaurs and plesiosaurs in the Mocuio Formation, higher in Chron C32n2n at 72.8 Ma, is 13.9°C, reflecting a relatively rapid drop in temperature. The mosasaurine-dominated Bench 19 Fauna was recovered from sediments dated to Chron C32n1n, about 71.5 Ma, which directly overlies a sampled interval that indicate temperatures of ∼18.5°C.
Examination of the δ13C curve presented in Strganac et al. (Reference Strganac, Jacobs, Ferguson, Polcyn, Mateus, Schulp, Morais, Tavares and Gonçalves2014a) shows declining, but relatively stable, values during deposition of the Baba Formation, but within the lower part of the Mocuio Formation a sharp negative excursion of ∼3‰ in δ13C values occurs with a recovery of equal magnitude coincident with the onset of the bench-forming depositional regime. This negative excursion is correlated with global patterns interpreted as the Campanian–Maastrichtian Boundary Events (tie points 9a and 8 sensu Voigt et al., Reference Voigt, Gale, Jung and Jenkyns2012), but the precipitous recovery (tie point 7), coincident with the base of Bench 19, appears strongest in the South Atlantic Ocean observed in foraminifera from the Walvis Ridge and Bentiaba, and thus may reflect increased localised primary productivity for this time, which may account at least in part for the large number of marine amniotes in this part of the section.
Mosasaurs originated in relatively low latitudes during the high temperatures of the Cenomanian (Polcyn et al., Reference Polcyn, Tchernov and Jacobs1999; Jacobs et al., Reference Jacobs, Ferguson, Polcyn and Rennison2005a,Reference Jacobs, Polcyn, Taylor and Fergusonb; Bardet et al., Reference Bardet, Houssaye, Rage and Superbiola2008). Before the end of the Cretaceous they occupied waters from the Antarctic to the tropics and into the northern high temperate latitudes. Their dispersal across the globe is accompanied by an increase in size and morphological disparity (Polcyn et al., Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014), and by the acquisition and maintenance of high body temperature (Bernard et al., Reference Bernard, Lécuyer, Vincent, Amiot, Bardet, Buffetaut, Cuny, Fourel, Martineau, Mazin and Prieur2010), reasonably considered in this case as an adaptation to colder water temperatures, especially in high latitudes. In addition, skin pigmentation preserved in exceptional specimens indicates dark colouration, which has been suggested to be an aid in thermoregulation (Lindgren et al., Reference Lindgren, Sjövall, Carney, Uvdal, Gren, Dyke, Schultz, Shawkey, Barnes and Polcyn2014). In this study we have seen temperature decrease in waters throughout most of the Late Cretaceous Period in one region along the Angola coast continuously inhabited by mosasaurs.
Acknowledgements
This publication results from Projecto PaleoAngola, an international cooperative research effort among the contributing authors and their institutions, funded by the National Geographic Society, the Petroleum Research Fund of the American Chemical Society, Sonangol EP, Esso Angola, Fundação Vida of Angola, LS Films, Maersk, Damco, Safmarine, ISEM at SMU, the Royal Dutch Embassy in Luanda, TAP Airlines, Royal Dutch Airlines and the Saurus Institute. We thank Margarida Ventura and André Buta Neto for providing our team with help in the field. Tako and Henriette Koning provided valuable support and friendship in Angola.




