


Introduction
Gebel Maghara is a dome-like structure covering ~400 km2 in the northern Sinai, 50 km south of the Mediterranean coast (Fig. 1). It comprises the thickest and most complete Jurassic outcrop in northern Sinai (1800 m; Al Far, Reference Al Far1966; Keeley, Reference Keeley1994). The Middle–Upper Jurassic succession of Gebel Maghara is well known for its rich and diverse macrobenthic fauna such as brachiopods, bivalves, gastropods, ammonites, and corals. This fauna has been repeatedly studied from a taxonomic point of view (e.g., brachiopods: Farag, Reference Farag1957, Reference Farag1959; Farag and Gatinaud, Reference Farag and Gatinaud1960a, Reference Farag and Gatinaudb; Feldman, Reference Feldman1987; Hegab, Reference Hegab1989, Reference Hegab1991; Feldman et al., Reference Feldman, Owen and Hirsch1991, Reference Feldman, Radulović, Hegab and Radulović2012; bivalves and gastropods: Douvillé, Reference Douvillé1916, Reference Douvillé1925; Hirsch, Reference Hirsch1980; and ammonites: Arkell et al., Reference Arkell, Bramkamp and Steineke1952; Parnes, Reference Parnes1988). The most-recent studies dealing with the stratigraphy and paleoecology are given by Abdelhady (Reference Abdelhady2014) and Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014, Reference Abdelhady and Fürsich2015a, Reference Abdelhady and Fürsich.b, Reference Abdelhady and Fürsichc).

Figure 1 Locality map.
The purpose of this paper is: (1) to describe the new bivalve taxon Costinuculana magharensis n. gen. n. sp. (order Nuculanida) from the Bathonian of Gebel Maghara and to compare it with similar taxa of the family Nuculidae; (2) to interpret the function of its oblique ribs, which cover an area up to ~45% of total valve length from the posterior end and to reconstruct its autecology; and (3) to determine its paleoenvironmental setting on the basis of facies relationships and associated faunal elements.
Geologic setting
The Jurassic succession of Gebel Maghara represents a sequential development of continental and marine sediments, starting in the Early Jurassic (Toarcian) and lasting until the Kimmeridgian (Fig. 2). It represents a wide variety of continental, deltaic, nearshore-siliciclastic, and carbonate-shelf environments (Al Far, Reference Al Far1966; Picard and Hirsch, Reference Picard and Hirsch1987), and contains a rich macrofauna. The marine strata are represented by the Rajabiah, Bir Maghara, and Masajid formations, whereas the continental sediments include the Mashabba, Shusha, and Safa formations. The nuculanid specimens have been collected from the middle to upper Bathonian Kehailia Formation (Fig. 2). This formation represents the lower member of Al Far’s (Reference Al Far1966) Masajid Formation (Kehailia Member). The Kehailia Formation unconformably overlies the lower Bathonian Safa Formation and underlies the Callovian Arousiah Formation (Fig. 2). It consists of yellow, highly fossiliferous marly silts and marls with thin, yellow glauconitic limestone intercalations and occasional sandstone interbeds. For more details about litho- and biostratigraphy see Abdelhady (Reference Abdelhady2014) and Abdelhady and Fürsich (Reference Abdelhady and Fürsich2015a, Reference Abdelhady and Fürsich.b, Reference Abdelhady and Fürsichc).

Figure 2 Chronostratigraphic correction chart of the Jurassic formations of Gebel Maghara, Sinai, Egypt (modified after Abdelhady and Fürsich, Reference Abdelhady and Fürsich2015a).
Materials and methods
The present study is based on 10 specimens with preserved shells that were collected from the middle to upper Bathonian Kehailia Formation. The preservation of the specimens is of varying quality; especially the posterior ends are mostly broken off. Except for a single left valve, they are preserved as articulated shells. They were prepared mechanically and carefully cleaned. For photography, they were whitened with magnesium oxide to accentuate finer ornamental details.
Linear measurements (taken with a Vernier caliper) are in millimeters. Orientation of measurements is shown in Figure 3.1 and 3.2. Abbreviations for dimensions are as follows: L=length, H=height, I=inflation of articulated specimens, D=distance from the anterior end to umbo (umbonal distance), nr=number of ribs along rostrum.

Figure 3 Measured parameters and orientation of Costinuculana magharensis n. gen. n. sp.: (1) lateral view of right valve; (2) dorsal view of articulated valves. Abbreviations: L=length, H=height, I=inflation of articulated specimen, D=distance from the anterior end to umbo, nr. ribs=number of oblique ribs along rostrum, RV=right valve, LV=left valve.
Repositories and institutional abbreviations
The specimens are housed in the collections of the Bayerische Staatssammlung für Paläontologie und Geologie in Munich under the prefix BSPG 2014 V.
Systematic paleontology
The systematic classification of the bivalves follows that of Carter et al. (2011). The morphological terminology follows the glossary of Cox (1969) in the Treatise on Invertebrate Paleontology.
Class Bivalvia Linnaeus, Reference Linnaeus1758
Subclass Protobranchia Pelseneer, Reference Pelseneer1889 (=Palaeotaxodonta Korobkov, Reference Korobkov1954)
Superorder Nuculaniformii Carter, Campbell, and Campbell, Reference Carter, Campbell and Campbell2000
Order Nuculanida Carter, Campbell, and Campbell, Reference Carter, Campbell and Campbell2000
Superfamily Nuculanoidea Adams and Adams, Reference Adams and Adams1858 (Gray, Reference Gray1854)
Family Nuculanidae Adams and Adams, Reference Adams and Adams1858 (Gray, Reference Gray1854)
Subfamily Veteranellinae Chen, Liu and Lan, Reference Chen, Liu and Lan1983
Remarks
Chen et al. (Reference Chen, Liu and Lan1983) relocated eleven nuculanid genera, including two new genera, to the new subfamily Veteranellinae. These genera are Veteranella Patte, Reference Patte1926; Glyptoleda Fletcher, Reference Fletcher1945; Nucundata Waterhouse, Reference Waterhouse1965; Girtyana Elias, Reference Elias1956; Teinonuculana Zhang in Zhang, Wang, and Zhou, Reference Zhang, Wang and Zhou1977; Ledoides Chen, Wen, and Lan in Gu et al., Reference Gu1976; Xiaoshuiculana Chen, in Chen, Liu, and Lan, Reference Chen, Liu and Lan1983; Eleganuculana Chen and Yang, Reference Chen and Yang1983; Indoculana Kanjilal and Singh, Reference Kanjilal and Singh1973; Costatoleda Roth von Telegd, Reference Roth von Telegd1914; Hilgardia Harris and Palmer, 1946; Qiongzhounia Lan in Chen, Liu, and Lan, Reference Chen, Liu and Lan1983; and Zealeda Marwick, Reference Marwick1924. These taxa are all strongly ornamented with one or more type of ribs (concentric, radial, and/or oblique). Based on the latter important taxonomic character, we place the new genus Costinuculana in the subfamily Veteranellinae. For more details and comparison among some of the above-mentioned genera, see below.
Genus Costinuculana new genus
Type species
Costinuculana magharensis n. gen. n. sp. from the middle Bathonian (Middle Jurassic) of Gebel Maghara, North Sinai, Egypt.
Diagnosis
Small, elongated, rostrate nuculanid, with posteriorly directed umbones. Rostrum with well-developed oblique ribs, variable in shape and size, asymmetrical in cross-section, straight to convex posteriorly, bifurcated ventrally and occasionally postero-dorsally, separated by wide and deep interspaces; escutcheon wide, moderately deep, and well ornamented with fine riblets, delimited by blunt ridges running from umbones to postero-dorsal margin; lunule narrow, lanceolate, sharply demarcated.
Etymology
Combination of costa (Latin)=rib, and the genus Nuculana, referring to the oblique ribs along rostrum.
Occurrence
The material of the new genus has been collected from the middle Bathonian ammonite Clydomphalites clydocromphalus Zone.
Remarks
The significant taxonomic characters of Costinuculana n. gen. are not present in the majority of Jurassic nuculanid species that have been identified and described from different localities (e.g., Pandey and Singh, Reference Pandey and Singh1981; Fürsich, Reference Fürsich1982; Aberhan, Reference Aberhan1994, Reference Aberhan1998; Jaitly et al., Reference Jaitly, Fürsich and Heinze1995; Sha et al., Reference Sha, Fürsich, Smith and Wang1998). The main diagnostic feature distinguishing Costinuculana from other genera of the family Nuculanidae (e.g., Nuculana Link, Reference Link1807) is the presence of well-developed oblique ribs on the posterior part of the flank, which cover an area up to ~45% of total valve length from posterior end (Fig. 4.1–4.9). The genus Ryderia Wilton, Reference Wilton1830 (type species: Leda renevieri Oppel, Reference Oppel1856, p. 215) is similar to the new genus in having an elongated-rostrate shell, a strongly rounded anterior margin, and small and opisthogyrate beaks, but differs in having a narrower and much more elongated rostrum, a narrow and smooth escutcheon, fine and closely spaced commarginal ribs, in lacking posterior oblique ribs, and in being much more compressed than the present material. The commarginal ribs of Glyptoleda Fletcher, Reference Fletcher1945 are folded and sub-vertically deflected towards the posterior part of the flank, but the ornamentation style completely differs from that of Costinuculana. In addition, Glyptoleda differs in being much larger than the present genus (L=~45 mm as opposed to 12.35 mm on average for C. magharensis n. sp.) and is of Permian origin. Chen et al. (Reference Chen, Liu and Lan1983) erected two genera, Xiaoshuiculana Chen in Chen et al. and Qiongzhounia Lan in Chen et al. from the Upper Triassic and Pliocene of China, respectively. The genus Xiaoshuiculana differs from Costinuculana n. gen. in having only radial ribs that cover the entire valves and in being shorter. The genus Qiongzhounia resembles Costinuculana n. gen. in having an elongated-rostrate shell and a well-developed umbonal posterior ridge, but differs also in the style of ornamentation. It is ornamented with faint concentric ribs on the flank and with delicate, weak growth lines on the posterior slope (see Chen et al., Reference Chen, Liu and Lan1983, p. 619, fig. 1). The reticulate ornamentation and short rostrum distinguishes Indoculana Kanjilal and Singh (Reference Kanjilal and Singh1973) from the Callovian of the Kachchh Basin, India, from Costinuculana n. gen. Ledoides Wen and Lan in Gu et al., Reference Gu1976 from the Upper Triassic of China differs in having three different kinds of ribs: concentric ribs on the umbonal area, a reticulate pattern on the anterior part, and oblique radial ribs on the posterior part. In addition, Ledoides has a narrow and shorter rostrum. The subgenus (Gonionucula) Hautmann and Aghababalou in Hautmann et al. (Reference Hautmann, Aghababalou and Krystyn2011, p. 22–24, figs. 2, 3) from the Norian–Rhaetian (Late Triassic) of Iran differs in having chevron-like ribs on the central part of the flank and an additional set of steeply opisthocline to nearly vertical ribs arranged in a narrow stripe on the posterior part of the flank. The latter authors stated that their new subgenus differs from the genus Acila Adams and Adams, Reference Adams and Adams1858 in having an additional set of opisthocline ribs on the posterior part. Apart from its ornamentation, the genus Acila differs from Costinuculana n. gen. in being less elongated (not rostrate), larger, and in having a wide and strongly rounded anterior margin.

Figure 4 Modified photographs (1, 2, 5–7, 9) and sketches (3, 8) showing the variation of opisthocline ribs of articulated specimens of Costinuculana magharensis n. gen. n. sp. from the middle to upper Bathonian Kehailia Formation of Gebel Maghara: (1, 2, 4, 7) side view of left valves showing straight and arched ribs with rounded crests; (1) BSPG2014V I3b; (2) BSPG2014V I3a; (4) BSPG2014V I4b; (7) BSPG2014V 1, holotype; (3, 6, 9) right valves showing bifurcated ribs close to the ventral and postero-dorsal margin; (3) BSPG2014V I3a; (6) BSPG2014V I4b; (9) BSPG2014V 1; (5) dorsal view showing the oblique angle between ribs and postero-dorsal margin; BSPG2014V I4b; (8) sketch of posterior part of Figure 4.7 showing the irregular ribs, bifurcated close to the ventral margin and occasionally towards postero-dorsal margin. Scale bar=2 mm.
Pandey and Singh (Reference Pandey and Singh1981, p. 1296, text-fig. 1) erected a new species “Indoculana” sadharaensis from the lower Callovian of Gora Dongar, India on the basis of the presence of faint radial riblets along the posterior part of flank. Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, p. 160) regarded the different types of surface ornament just as intraspecific variations of Nuculana (Praesaccella) calloviensis (Kanjilal and Singh, Reference Kanjilal and Singh1973). The second feature regarded by Pandey and Singh (Reference Pandey and Singh1981) as diagnostic feature of “I.” sadharaensis, is the presence of transverse threads in the escutcheon. Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, p. 160, pl. 2, figs. 13, 14) observed the same transverse threads in N. (P.) calloviensis, which are therefore of no specific significance. In addition, “I.” sadharaensis differs from C. magharensis n. sp. in having a rapidly narrowing and short rostrum, less-developed postero-dorsal ridge, faint radial and commarginal ribs, and in being less inflated. N. (P.) calloviensis (Kanjilal and Singh, Reference Kanjilal and Singh1973), as figured and described by Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995), from the Callovian of Kachchh differs in having a reticulate ornamentation, which covers the entire valve, a less-elongated rostrum, and in being less inflated and smaller than the present species. The valve of Ryderia texturata (Terquem and Piette, Reference Terquem and Piette1865), described and figured by Hodges (Reference Hodges2000, p. 45, pl. 3, figs. 5, 8–11, 13–17) and Aberhan et al. (Reference Aberhan, Scholz and Schubert2011, p. 70, pl. 1, fig. 7) from the Lower Jurassic of southwestern England and the Herforder Liasmulde, Germany, respectively, carries faint vertical threads in the posterior part of the flank, which start slightly below the postero-dorsal ridge and do not reach the ventral margin. The ribbing style of C. magharensis n. sp. is, indeed, completely different from that of the German material. In addition, R. texturata differs in having a very narrow and strongly elongated rostrum, strongly convex anterior and ventral margins, and in being larger (L=~42 mm as opposed to 14.3 mm on average) and quite compressed.
Hautmann et al. (Reference Hautmann, Aghababalou and Krystyn2011, p. 22, figs. 2–4) erected the new subgenus and species Trigonucula (Gonionucula) aciloides from the Upper Triassic of Iran based on oblique ribs along the posterior flank. Their species differs from the Egyptian material by its more complex ornamentation pattern. The ribs are chevron-like on the central part of the flank and an additional set of steeply opisthocline to nearly vertical ribs are arranged in a narrow strip on the posterior part of the flank. Apart from the ribbing style, Trigonucula (G.) aciloides differs in having less-elongated valves, a smooth escutcheon, more or less central umbones, slightly rostrate valves, and in being less inflated. Veteranella (Ledoides) langnongensis Wen and Lan in Gu et al., Reference Gu1976, as figured and described by Fang et al. (Reference Fang, Chen, Chen, Sha, Lan and Wen2009, p. 12, fig. 3.1–3.5), from the Upper Triassic of China differs in having a greater number of oblique to sub-vertical ribs along the posterior flank, a reticulate ornamentation on the central and anterior parts (except the umbonal area), a slightly convex ventral margin, sharp beaks, a narrower and longer rostrum, and in being much more elongated than the present species.
Costinuculana magharensis new species

Figure 5 Costinuculana magharensis n. gen. n. sp. from the middle-upper Bathonian Kehailia Formation of Gebel Maghara, Egypt. All specimens are in shell preservation and articulated except Figure 5.21. In all specimens the tip of the rostrum is broken off: (1) right valve view; (2) dorsal view; BSPG2014V I4a; (3) left valve view showing thick irregular ribs with wide and deep interspaces; (4) right valve view with well-developed commarginal ribs; BSPG2014V I3b; (5) left valve view showing thick irregular ribs with sharp crests; (6) dorsal view; (7) right valve view with bifurcated ribs close to the ventral margin; BSPG2014V I4b; (8) right valve view; (9) close-up showing an inverted V-shaped rib close to the ventral margin; BSPG2014V I4c; (10) left valve view showing slightly folded ribs posteriorly and bifurcated ventrally; (11) right valve view; (12) dorsal view showing the lanceolate and demarcated lunule; BSPG2014V I3a; (13) left valve view, holotype; (14) close-up of Figure 5.13 showing the irregular and bifurcated ribs close to the ventral margin, (15) right valve view with serpulid; (16) dorsal view showing the moderately inflated valves, (17) close-up of Figure 5.16 showing wide and ornamented escutcheon, delimited by blunt ridges; BSPG2014V 1; (18) right valve view showing thick and well-developed commarginal ribs close to the ventral margin; (19) dorsal view; (20) left valve view; BSPG2014V I2: (21) right valve view showing numerous chevron-shaped teeth; BSPG2014V I5c. Scale bars=2 mm.
2014 Nuculoidea n. sp. Reference AbdelhadyAbdelhady, p. 172, fig. 5.4A, 5.4B.
2014 Nuculoidea n. sp.; Reference Abdelhady and FürsichAbdelhady and Fürsich, p. 181, fig. 6A, 6B.
Holotype
Articulated specimen no. BSPG 2014V 1 (Fig. 5.13–5.17) from the middle Bathonian (Middle Jurassic) of Gebel Maghara, Egypt.
Paratypes
Eight articulated specimens no BSPG 2014V I2, I3a, b, I4a, b, I5a, b from the middle Bathonian of Gebel Maghara, and one right valve no BSPG 2014V I5c.
Type locality
Gebel Maghara, North Sinai, Egypt.
Diagnosis
See genus.
Description
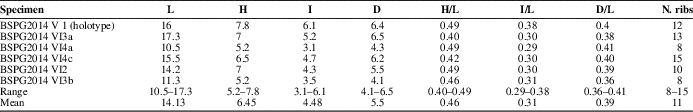
Shells small (L=10.5–17.3 mm; Table 1), elongated-rostrate in outline, strongly inequilateral, equivalved, moderately inflated (I/L=29–38%), and posteriorly elongated with ribbed rostrum (Fig. 4). Antero-dorsal margin short, strongly convex, higher than postero-dorsal one, and meeting anterior margin in strongly rounded curve. Postero-dorsal margin long, slightly concave. Anterior margin strongly convex, meeting ventral margin in rounded angle. Posterior margin rostrate, its end broken off. Ventral margin broad, irregular, moderately convex toward anterior margin, slightly concave posteriorly, and forming rostrum with postero-dorsal margin. Umbones broad and strongly convex. Beaks small, strongly incurved posteriorly and situated, on average, 39% of total shell length from anterior end (Table 1). Escutcheon wide, moderately deep, ornamented with fine riblets, and delimited by blunt ridge; running from umbo to postero-dorsal margin (Figs. 4.5, 5.16, 5.17). Lunule narrow, lanceolate, deep, and demarcated (Fig. 5.12, 5.19). Hinge with numerous chevron-shaped teeth, points directed towards umbones (Figs. 5.21, 6). Anterior hinge carrying slightly more teeth (14) than posterior hinge (≥11). Ornamentation consisting of numerous well-developed commarginal ribs on anterior and central parts of valve, but irregular and opisthocline ribs along rostrum (9–12), separated by wide and deep interspaces (Fig. 4.1–4.9). Commarginal ribs well developed ventrally (Fig. 5.18), asymmetrical in cross-section with slightly convex, broad ventral sides and steeply inclined and slightly concave dorsal sides (Fig. 7.2). Oblique ribs variable in shape and size, straight to folded, bifurcated ventrally and occasionally postero-dorsally (Figs. 4.3, 4.4, 4.6–4.8, 5.7–5.9, 5.13, 5.14), covering an area ~45% of total valve length from posterior end. Close to ventral margin, these ribs are thicker, bifurcating, and are asymmetrical in cross-section with a steeply concave side anteriorly and slightly convex side in opposite direction (Fig. 7.2).

Figure 6 Hinge of the right valve of Costinuculana magharensis n. gen. n. sp. from the middle to upper Bathonian Kehailia Formation of Gebel Maghara; BSPG2014V I5c.

Figure 7 (1) Reconstruction of the Nicaniella (N.) pisiformis association of which Costinuculana magharensis n. gen. n. sp. is a member. 1=Nicaniella (N.) pisiformis J. de C. Sowerby; 2=Ryderia decorata (Douvillé); 3=Costinuculana magharensis n. gen. n. sp. 4=Palaeonucula lateralis (Terquem and Jourdy); 5=Pholadomya (Ph.) inornata (J. de C. Sowerby); 6=Nuculoma variabilis (J. de C. Sowerby); 7=Nanogyra nana (J. Sowerby); 8=Daghanirhynchia angulocostata Cooper; 9=Gervillella orientalis (Douvillé); 10=Amphitrochus magharensis, Douvillé; 11=Africogryphaea costellata (J. de C. Sowerby); 12=Thalassinoides; (2) mode of life of Costinuculana magharensis. is=inhalant siphon; es=exhalent siphon; pp. palp proboscides; t=tentacle. While the commarginal ribs aid the burrowing process, the oblique posterior ribs stabilize the shell within the substrate; (3) relative abundance (%) of some species of the trophic nucleus of the Nicaniella (N.) pisiformis association (number denotes taxa in Figure 7.1). Data taken from Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014).
Table 1 Measurements (in mm) of Costinuculana magharensis n. gen. n. sp. from the Middle Bathonian of Gebel Maghara, Sinai. Abbreviations given in Figure 3.
Etymology
After the type locality Gebel Maghara, Sinai Peninsula, Egypt.
Remarks
See genus.
Mode of life and functional morphology of Costinuculana n. gen.
Morphologically, Costinuculana n. gen. is narrowly elongated, rostrate, has a broadly convex anterior margin, and is moderately inflated with strong, oblique ribs along the rostrum. The long narrow valve (like in Ryderia) indicates a rapid burrower (Stanley, Reference Stanley1970). Recent nuculanids are shallow infaunal bivalves with inhalant and exhalant siphons, which usually burrow with their long axis vertical or steeply inclined (Cox, Reference Cox1960). The posterior end is maintained above the sediment-water interface and the posterior siphons are never covered with sediment. They are deposit-feeders, feeding from the sediment surface by palp proboscides (Cox, Reference Cox1960). These palp proboscides extend immediately ventral of the siphons and collect food from the surface of the substrate (Yonge, Reference Yonge1939, p. 86).
With respect to ribbing style, most ribs in bivalves are commarginal (“concentric”) or radial (longitudinal) (Checa and Jiménez-Jiménez, Reference Checa and Jiménez-Jiménez2003). In a third, less-common ribbing pattern, the ribs are oblique (Seilacher, Reference Seilacher1972) or discordant (Stanley, Reference Stanley1969), whereby ribs migrate along the margin with growth. According to Stanley (Reference Stanley1969, Reference Stanley1970) and Seilacher (Reference Seilacher1972, Reference Seilacher1973), one of the functions of oblique ribs in bivalves is to facilitate burrowing in soft sediments. For a burrowing-related function, ribs show a gentle slope in the burrowing direction and a steep, in some cases inverted, slope in the opposite direction (frictional asymmetry; Seilacher, Reference Seilacher1972). For some other cases, Seilacher (Reference Seilacher1972) assumed that the oblique ribs were non-functional or had functions unrelated to burrowing. Later, Stanley (Reference Stanley1981) and Savazzi (Reference Savazzi1982, Reference Savazzi1983) showed experimentally that the radial ribs of several cardiids and of one arcid act to reduce the number of burrowing sequences required to complete burrowing.
Costinuculana magharensis n. gen. n. sp. has two kinds of ribs, commarginal and oblique ribs. The commarginal ribs are asymmetrical in cross-section, with a wide and slightly convex side in the burrowing direction, and a steeply inclined and slightly concave side in the opposite direction (Fig. 7.2). In contrast, the oblique ribs are thick and asymmetrical in cross-section close to the ventral margin, with a steeply concave side in the burrowing direction and a slightly convex side in the opposite direction (Fig. 7.2). At the beginning of burrowing, when the bivalve erected itself, the asymmetrical commarginal ribs facilitated the burrowing process (burrowing-related function). Once most of the bivalve had buried itself, the posterior oblique ribs provided stability for the organism in the soft substrate when the ribs were nearly perpendicular to the direction of burrowing (Fig. 7.2). In this case, the concave sides of the oblique ribs caught the sediment when the shell rotated forward during the burrowing process and kept the posterior end above the sediment-water interface. Because the posterior end of the shell presumably remained above the sediment-water interface, the thick posterior-most oblique ribs probably had another function. Most likely, they strengthened the thin and elongated rostrum against durophagous predators such as ammonites. The inflated valves with their broadly convex anterior margin in addition helped the bivalve to maintain a stable position within the soft substrate and to keep the posterior end above the sediment-water interface.
Paleosynecological remarks
Based on the cluster analysis of Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014, p. 179, fig. 3), Costinuculana magharensis n. gen. n. sp. is a member of their Nicaniella (N.) pisiformis association. This association is found mostly in fine-grained sediments (mainly marly silt and marl) of the middle to upper Bathonian Kehailia Formation of Gebel Maghara. The dominant organisms of this association are bivalves (75.4%), whereas gastropods, brachiopods, corals, and echinoids are subordinate. According to Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014), the trophic nucleus consists of 12 species and is dominated by small, shallow-burrowing Nicaniella (N.) pisiformis J. de C. Sowerby (31.5%; Fig. 7.3), Nuculoma variabilis (J. de C. Sowerby) (10.9%), and Ryderia decorata (Douvillé) (8.3%). Shallow-infaunal organisms represent 67.8% of the total benthic fauna followed by epifaunal ones 28.2% (Fig. 7.1). With respect to the feeding habits, suspension-feeders dominate (54.2%), followed by deposit-feeders (28.1%), omnivores (12.7%), and microcarnivores (4.9%).
The fine-grained sediment and the abundance of shallow-infaunal organisms and deep-burrowing bivalves (altogether 67.8%) indicate that the Nicaniella (N.) pisiformis association occupied a soft substrate. Due to the activities of mobile taxa (40.7%; Abdelhady and Fürsich, Reference Abdelhady and Fürsich2014), the softness of substrate probably increased and thus became less attractive for epifaunal organisms. The Nicaniella (N.) pisiformis association reflects low-energy conditions, in which physical breakage was insignificant. All specimens of Costinuculana n. gen. except one are articulated, which implies a relatively low degree of reworking. According to the high percentage of suspension-feeders (54.2%), water energy must have been sufficient enough to keep nutrients in suspension. Moreover, the high diversity both in terms of evenness and species richness and the presence of stenohaline groups, such as ammonites and echinoids, indicate normal-marine, fully oxygenated low-stress conditions. For more details about the paleo-community see Abdelhady (Reference Abdelhady2014, p. 170) and Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014, p. 179–184).
Conclusions
-
(1) The bivalve Costinuculana magharensis n. gen. n. sp. (Nuculanida, Nuculanidae) is described from the middle to upper Bathonian Kehailia Formation of Gebel Maghara, North Sinai, Egypt.
-
(2) The main morphological feature that distinguishes Costinuculana n. gen. from other nuculanid genera is the irregular opisthocline ribs on the rostrum, which cover an area ~45% of the total valve length from the posterior end. Other diagnostic features are the morphology of the escutcheon and lunule.
-
(3) Costinuculana magharensis n. gen. n. sp. differs from related species by its small size and its well-developed irregular opisthocline ribs on the rostrum, moderately inflated valves, narrow, lanceolate, and sharply demarcated lunule, and wide and deep escutcheon with well-developed riblets.
-
(4) The conspicuous oblique ribs in C. magharensis n. sp. probably helped the bivalve to maintain a stable position in the soft substrate. The oblique and bifurcated ribs increased the friction between the valves and sediment. In addition, these ribs probably reinforced the thin, elongated rostrum against durophagous predators (e.g., ammonites).
-
(5) Paleosynecologically, the Nicaniella (N.) pisiformis association and the fine-grained substrate reflect low-energy conditions. The species diversity and evenness of the association indicate a stable environment suitable for the colonization by several guilds.
Acknowledgments
The authors greatly appreciate the thoughtful reviews and valuable suggestions by Associate Editor M. Hautmann and two anonymous reviewers. A.A.A. wants to thank the German Academic Exchange Service (DAAD) and the Egyptian Mission (Ministry of High Education of Egypt) for their financial support through the German-Egyptian Research Long-term Scholarship (GERLS).








