


Introduction
Hybridization has been used as one of the range of biotechnology tools in solving many aquaculture problems (Rahman et al., Reference Rahman, Arshad, Marimuthu, Ara and Amin2013), the mating of genetically differentiated individuals or groups, (i.e. crosses within a species or between different species) (Bartley et al., Reference Bartley, Rena and Immink2001). Hence, it is able to transfer desirable traits from one group/species to another (Rahman et al., Reference Rahman, Uehara and Aslan2000, Reference Rahman, Uehara and Lawrence2005). The desirable trait may include, but not be limited to, an increase in growth rate, improved productivity, better fillet quality, disease resistance, increase in environmental tolerance, better food conversion, sexual dimorphism and increased harvesting rate in culture systems (Rahman et al., Reference Rahman, Arshad, Marimuthu, Ara and Amin2013).
Pangasianodon hypophthalmus (Sauvage, 1878) and Clarias gariepinus (Burchell, 1822) are among the most emblematic and important freshwater aquaculture species in south-east Asia, West Africa and many parts of the world (Hung et al., Reference Hung, Tam, Cacot and Lazard1999; Solomon et al., Reference Solomon, Okomoda and Ochai2013). P. hypophthalmus has a better fillet quality and can grow to a very large size (Hung et al., Reference Hung, Tam, Cacot and Lazard1999; Chattopadhyay et al., Reference Chattopadhyay, Mazumder and Mazumdar2002). The artificial breeding pattern involves stripping both male and female, however transient mortality during the first feeding days is a major challenge in the larva nursing of this catfish (Baras et al., Reference Baras, Slembrouck, Cochet, Caruso and Legendre2010). It also has a late maturity period which is between 2−3 years (Hung et al., Reference Hung, Tam, Cacot and Lazard1999). C. gariepinus, however, has a faster growth and matures early (9 months). The possession of a suprabranchial organ as opposed to the vascularized swim bladder found in P. hypophthalmus makes the African catfish more tolerant to anoxic water than its Asian counterpart (Ahmed et al., Reference Ahmed, Mohamed, Ahmed and Masoud2008). However, the gonad morphology of the male C. gariepinus makes sperm collection by stripping impossible. Hence, artificial breeding procedure involves sacrificing the male to obtain the sperm. This breeding system limits the possibility of stock improvement by selective breeding (Legendre et al., Reference Legendre, Teugels, Cauty and Jalabert1992). Furthermore, the high rate of cannibalism due to social interaction in the African catfish can significantly reduce the number of fish at harvest (Almaza´n Rueda, Reference Almaza´n Rueda2004; Olufeagba & Okomoda, Reference Olufeagba and Okomoda2016).
Intergeneric and interspecific hybridization of P. hypophthalmus and C. gariepinus with many close species has been successfully carried out, some of which showed positive heterosis for growth, survival and many other desirable traits (Tober et al., Reference Tober, Gabriele and Hans-Jurgen1995; Sahoo et al. Reference Sahoo, Giri, Sahu and Ayyappan2003; Gustiano Reference Gustiano2004; Hassan et al., Reference Hassan, Azmi and Putra2011; Olufeagba et al., Reference Olufeagba, Okomoda and Shaibu2016). Earlier studies by Boonbrahm et al. (Reference Boonbrahm, Tarnchalanukit and Suraniranat1977), Tarnchalanukit (Reference Tarnchalanukit1985, Reference Tarnchalanukit1986) and Na-Nakorn et al. (Reference Na-Nakorn and Rangsin1993) had also demonstrated successful hybridization between some species of the Clariid family [Philippine catfish Clarias batrachus (Linnaeus, 1758) and Bighead catfish Clarias macrocephalus Gunther, 1864] with the Asian catfish P. hypophthalmus. However, successfully hybridization between P. hypophthalmus and C. gariepinus has not been scientifically reported to date. Successful hybridization of these fish could offer solutions to some of the problems associated with the breeding of both pure species. For instance, the killing of male C. gariepinus to obtain testis is eliminated due to the ease of sperm stripping from the male P. hypophthalmus. Also, the early maturity and high fecundity of the female C. gariepinus complements the difficulty faced with the female P. hypophthalmus brood fish. The present study therefore analyses for the very first time the breeding performance, survival and growth of the hybrids and pure crosses of these two species during the embryonic and larval periods.
Materials and Methods
Brood fish procurement and breeding
Eight brood fish each of P. hypophthalmus (mean weight of 1.7 kg) and C. gariepinus (mean weight of 1 kg) of reproductive age (above 3 years and 1 year respectively) were obtained from the School of Fisheries and Aquaculture Sciences hatchery of the Universiti Malaysia Terengganu, in Malaysia (1:1 male to female). They were acclimatized for 2 weeks in rectangular fibreglass tanks and fed on a commercial diet (35% crude protein). In two breeding trials, hybridization was attempted between P. hypophthalmus and C. gariepinus using eight brood fish for each trial (i.e. two pairs of male and female for both species). Both sexes of P. hypophthalmus were first injected with Ovaprim® hormone at a rate of 0.2 ml and 0.5 ml of hormone per kg of the fish (for female and male respectively). Female P. hypophthalmus were given a second injection 8 h later at a dosage of 0.3 ml of Ovaprim® hormone per kg (to make up recommended dosage of 0.5 ml per kg) (Chaturvedi et al., Reference Chaturvedi, Ambulkar, Singh and Pandey2015). Female C. gariepinus were injected a one-time dosage of 0.5 ml Ovaprim® hormone per kg at the same time the second injection was administered to the female P. hypophthalmus. This procedure was aimed at synchronising the timing of ovulation and stripping for both species that had different latency period of 16 h and 8 h respectively for P. hypophthalmus and C. gariepinus. The fish were maintained in eight separate tanks according to their sex and species. Eggs from each female were stripped into two bowls according to their species. This was gently mixed and half of the eggs from each species transferred into another bowl to obtain four batches of eggs for the various directional crosses (comprising of two pure and two hybrid crosses). A small portion of the eggs (15−25 eggs) from both species was also isolated till they become opaque to determine fertilization rate. Milt from male P. hypophthalmus was obtained by stripping. The males C. gariepinus, however, were tranquilized with 150 mg/1 solutions of tricaine methane sulphonate (MS222) (Wagner et al., Reference Wagner, Jeppsen, Arndt, Routledge and Bradwisch1997) before they were sacrificed. The testes were macerated into a small bowl to mix the sperm content of both males (of C. gariepinus). Half of the content was used for the pure crosses while the other half was used for the hybrid crosses based on the direction shown below:
-
• ♀C. gariepinus × ♂C. gariepinus, (♀CG × ♂CG)
-
• ♀C. gariepinus × ♂P. hypophthalmus (♀CG × ♂PH)
-
• ♀P. hypophthalmus × ♂C. gariepinus, (♀PH × ♂CG)
-
• ♀P. hypophthalmus × ♂P. hypophthalmus, (♀PH × ♂PH).
The eggs and sperm content were mixed uniformly for 1 min, after which a small quantity of water (100 ml) was added and the content mixed again for another minute. The excess water and sperm were decanted leaving behind the fertilized eggs. Triplicate batches of equal eggs mass (10 g) were spawned on 12 nylon mesh substrate suspended over continuously oxygenated water in 12 aquarium bowls (0.5 × 0.5 × 0.5 m3). The aquaria were tagged appropriately in accordance to the crosses they represent. The numbers of egg in 1 g of fertilized egg mass were also determined for each cross to estimate the number of eggs spawned and determine associated breeding parameters.
Determination of breeding and growth performance
The time taken for the small portion of the eggs initially separated to become opaque (dead eggs) was noted to estimate fertilization rate using the formulae specified by Ella (1987) as shown below:
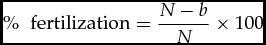 $$\begin{equation*}
\%\,\, {\textrm{ fertilization}} = \frac{{N - b}}{N} \times 100
\end{equation*}$$
$$\begin{equation*}
\%\,\, {\textrm{ fertilization}} = \frac{{N - b}}{N} \times 100
\end{equation*}$$
where (N) represents the total number of eggs spawned, (b) number of bad eggs and was obtained by counting.
The hatching rate of each cross was evaluated by expressing the value of hatch fry as a percentage of the total number of eggs incubated:
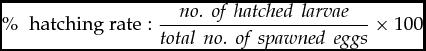 $$\begin{equation*}
\%\,\, {\textrm{ hatching rate}}:\frac{\textit{no.\, of\, hatched\, larvae}}{\textit{total\, no.\, of\, spawned\, eggs}} \times 100
\end{equation*}$$
$$\begin{equation*}
\%\,\, {\textrm{ hatching rate}}:\frac{\textit{no.\, of\, hatched\, larvae}}{\textit{total\, no.\, of\, spawned\, eggs}} \times 100
\end{equation*}$$
The number of normal and deformed larvae was also determined by direct observation and counting. Generally, the criteria used to determine normality of hybrid hatchlings were the presence of a straight body and a distinct head distinguished from the yolk. Divergence from this form was considered abnormal. Post-yolk absorption survival was estimated (at first feeding), 100 larvae from each cross were then stocked in 0.5 × 0.5 × 0.5 m3 aquarium tanks using a static system with continuous aeration (natural photoperiod of 12 h daylight and 12 h darkness). Each group was fed sequentially a dietary regime of live Artemia; fishmeal and commercial micro-pellets feed. Artemia was fed as the first diet between the 3rd to 21st days post hatching (dph). They were incubated and hatched in salt water 24 h prior to feeding. Each batch of hatched Artemia was fed continuously to the fish for a maximum of 3 days, after which newly hatched Artemia was used. Artemia was administered four times daily (between 8:00 h and 21:00 h). The fishmeal was only fed to the larvae on the 22nd dph (feeding was stopped due to increased mortality of the fish). The fishmeal was made into a dough and stocked to the side of the aquarium tanks for sequential release into the culture system, this was done three times a day. From the 23rd to the 35th dph commercial micro-pellets feed (45% crude protein) were distributed ad libitum by hand three times a day. Growth parameters of the hatchlings were observed under these feeding regimes. Fish were bulk weighed weekly using a sensitive weighing balance (nearest 0.00 mg) and mean weights obtained (Fig. 1). Total lengths of 15 randomly selected hatchlings were also taken (nearest 0.00 cm) at the start of the exogenous feeding and at the end of the 35 dph using a micrometre gauge.

Figure 1 Weekly growth of pure and reciprocal hybrids of Pangasianodon hypophthalmus and Clarias gariepinus.
All fish were returned to the appropriate rearing tank after measurements were taken. Mortality in the rearing tanks was noted daily and recorded appropriately. Also, dead fishes were observed for missing parts which are evidence of incomplete cannibalism (Almaza´n Rueda, Reference Almaza´n Rueda2004; Olufeagba & Okomoda, Reference Olufeagba and Okomoda2016). Upon weekly checks for the measurement of growth, survivors were counted and missing fish were assumed to have succumbed to complete cannibalism (Solomon & Okomoda, Reference Solomon and Okomoda2012; Olufeagba & Okomoda, Reference Olufeagba and Okomoda2016). Growth parameter determined in the fishes includes:
mean length gained (cm) = L2 − L1
mean weight gained (mg) = W2 − W1
growth rate (mg/day) =
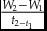 $\frac{{{W_2} - {W_1}}}{{{t_{2 - {t_1}}}}}$
$\frac{{{W_2} - {W_1}}}{{{t_{2 - {t_1}}}}}$
specific growth rate (%/day) =
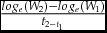 $\frac{{lo{g_e}( {{W_2}} ) - lo{g_e}( {{W_1}} )}}{{{t_{2 - {t_1}}}}}$
$\frac{{lo{g_e}( {{W_2}} ) - lo{g_e}( {{W_1}} )}}{{{t_{2 - {t_1}}}}}$
where W1 = initial weight (mg); W2 = final weight (mg); L1 = initial length (cm); L2 = final length (cm); t2 − t1= duration between W2 and W1(d).
survival rate (%) =
 $\frac{{{\rm{fish\ stocked}} - {\rm{mortality}}}}{{{\rm{\ fish\ stocked}}}}{\rm{\ }} \times {\rm{\ }}100$
$\frac{{{\rm{fish\ stocked}} - {\rm{mortality}}}}{{{\rm{\ fish\ stocked}}}}{\rm{\ }} \times {\rm{\ }}100$
Note: Survival was recorded both in cumulative terms (i.e. based on the number of larvae initially stocked in the aquarium at 3 dph) and in relative terms (i.e. based on the previous number of surviving fish in the culture tanks before the occurrence of the current mortality):
cannibalism mortality (%) =
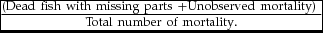 $ \frac{{( {{\rm{Dead\ fish\ with\ missing\ parts\ }} + {\rm{Unobserved\ mortality}}} ){\rm{\ }}}}{{{\rm{Total\ number\ of\ mortality}}.}}$
× 100
$ \frac{{( {{\rm{Dead\ fish\ with\ missing\ parts\ }} + {\rm{Unobserved\ mortality}}} ){\rm{\ }}}}{{{\rm{Total\ number\ of\ mortality}}.}}$
× 100
heterosis H (%) =
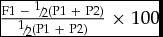 $\frac{{{\rm{F1\ -\ \raise.5ex\hbox{$\scriptstyle 1$}\kern-.1em/ \kern-.15em\lower.25ex\hbox{$\scriptstyle 2$} (P1\ + \ P2)}}}}{{{\rm{\raise.5ex\hbox{$\scriptstyle 1$}\kern-.1em/ \kern-.15em\lower.25ex\hbox{$\scriptstyle 2$} (P1\ + \ P2)}}}}{\rm{\ \times \ 100}}$
$\frac{{{\rm{F1\ -\ \raise.5ex\hbox{$\scriptstyle 1$}\kern-.1em/ \kern-.15em\lower.25ex\hbox{$\scriptstyle 2$} (P1\ + \ P2)}}}}{{{\rm{\raise.5ex\hbox{$\scriptstyle 1$}\kern-.1em/ \kern-.15em\lower.25ex\hbox{$\scriptstyle 2$} (P1\ + \ P2)}}}}{\rm{\ \times \ 100}}$
where, F1, P1, and P2 are the averages of the performance of the first generation of hybrids, parent 1 and parent 2, respectively.
At the end of the experiment, the gross morphologies of the surviving hybrids and pure species were compared. Morphological variation in the hybrids was described and assumed as different morphotypes. Their ratios based on the pooled surviving hybrids in all the rearing tanks were calculated and recorded morphotype were described based on the observable fin and body characteristic. Detailed morphological comparison could not be made at 35 dph due to the size of the fish.
Water quality parameter
Water quality parameters such as temperature, dissolved oxygen, total dissolved solids, pH, and ammonia concentration were monitored daily throughout this study using a YSI professional plus multi-parameter water quality meter (Model 13M10065, USA). Also, when mass mortality (≥50%) was noticed within 24 h in any of the group (between 3rd and 35th dph), water quality parameter during that period were separately and analysed to see if mortality could be linked to degradation of water quality. A 24 h challenge test was undertaken on the 36 dph to roughly evaluate the tolerance of the different crosses to low dissolved oxygen. During this challenge test, the surviving fishes from the previous study were raised under the same static condition with no aeration. Water quality parameters were recorded and survival determined.
Data analysis
Descriptive statistics were analysed using mini tab 14 computer software followed by one-way analysis of variance (ANOVA). When significant (P < 0.05) differences were observed, data were separated using Fisher's least significant difference.
Results
Breeding performance in this study is summarized in Table 1. Both pure and reciprocal hybrids had comparable (P > 0.05) fertilization rates (86–89%). Artificial propagation of the reciprocal hybrid was successful in producing viable larvae. However, hatchability was about four times as high for ♀CG × ♂PH than for ♀PH × ♂CG (49 vs 12%) but generally lower than values recorded in the pure sibling (88 and 76% for ♀CG × ♂CG and Panga respectively); 67.33 and 51.09% of ♀CG × ♂PH and ♀PH × ♂CG respectively were abnormal while pure siblings had abnormality less than 5%.
Table 1 Breeding parameters and heterosis (H) of performance for the reciprocal hybridization between Pangasianodon hypophthalmus and Clarias gariepinus

a,b,c Means in the same row with different superscript letters differ significantly (P ≤ 0.05).
At the end of the endogenous feeding, close to half of the hatched larvae of P. hypophthalmus (45.20%), ♀CG × ♂PH (50.33%) and ♀PH × ♂CG (45%) survived. Survival was much higher in pure Clarias (75%) compared with pure Panga (45.2%). During the Artemia feeding period (Table 2), survival was significantly higher in fish originating from Clarias eggs (82–87%) than in those originating from Panga eggs (49−56%) (Fig. 2). However, when fish were offered micro-pellets, survival was high in all groups, but proportionally lowers in pure Clarias and ♀CG × ♂PH (81–85 versus 91%). This was partly due to different rates of cannibalism, which accounted for over 95% of total mortality in pure Clarias and 69.3% in ♀CG × ♂PH as against ≤15% recorded in the other crosses.
Table 2 Survival of pure and reciprocal hybrids of Pangasianodon hypophthalmus and Clarias gariepinus under different feeding regimes

a,b,c Means in the same row with different superscript letters differ significantly (P ≤ 0.05).

Figure 2 Cumulative survival of pure and reciprocal hybrids of Pangasianodon hypophthalmus and Clarias gariepinus under different experimental conditions.
The 24-h challenge test (i.e. tolerance to low dissolved oxygen) at the end of the rearing period (Table 3) revealed a good hardiness of pure Clarias, pure Panga and ♀PH × ♂CG. They all exhibited survival rates over 90% during this test. By contrast, less than 10% of ♀CG × ♂PH survived this test. The greater fragility of ♀CG × ♂PH to degradation of water quality is supported by the lower survival rate that had been observed at 22 dph, at the transition between the two types of food: only 38% of the ♀CG × ♂PH survived this transition, whereas survival in the other crosses ranged from 89.5 to 96.5% (Table 2).
Table 3 Survival of pure and reciprocal hybrids of Pangasianodon hypophthalmus and Clarias gariepinus and the mean water quality of the experimental unit during the 24 h challenge test without aeration

a,b Means in the same row with different superscript letters differ significantly (P ≤ 0.05).
The performance of larvae after a 35 dph feeding is shown in Table 4. The final mean body weight varied from 833 to 436 mg and specific growth rate from 11.64 to 12.85% day−1. These parameters were significantly lower in P. hypophthalmus and ♀PH × ♂CG compared with the rest of the crosses. However, ♀CG × ♂PH hybrid displayed a 60.37% heterosis in mean body weight relative to the performance of the mid parents, while ♀PH × ♂CG had a negative heterosis (−16.49%).
Table 4 Growth parameters and heterosis (H) of performance for the reciprocal hybridization between Pangasianodon hypophthalmus and Clarias gariepinus

a,b,c Means in the same row with different superscript letters differ significantly (P ≤ 0.05).
Two morphotypes were observable in the ♀CG × ♂PH at 35 dph (Table 5). The first morphotype (87% of the hybrid pool) has combined features of both parents, while the other morphotype (13% of the hybrid pool) was hardly distinguished from the C. gariepinus parent. The ♀PH × ♂CG hybrid, however, has only one morphotype which looks closely and indistinguishable from the P. hypophthalmus parent.
Table 5 Morphological features of pure and reciprocal hybrids of Pangasianodon hypophthalmus and Clarias gariepinus larvae that survived up to 35 dph

Discussion
This study demonstrates the possibility of producing viable hybrids from reciprocal crosses of P. hypophthalmus and C. gariepinus. Tarnchalanukit (Reference Tarnchalanukit1986), and Boonbrahm et al. (Reference Boonbrahm, Tarnchalanukit and Suraniranat1977) had earlier reported successful hybridization between C. batrachus and P. hypophthalmus. Similarly, Na-Nakorn et al. (Reference Na-Nakorn and Rangsin1993) and Tarnchalanukit (Reference Tarnchalanukit1985) also reported their findings between P. hypophthalmus and C. macrocephalus. Glamuzina et al., (Reference Glamuzina, Tutman and Skaramuca1999, Reference Glamuzina, Glavić, Skaramuca, Kozul and Tutman2001). Frisch & Hobbs, (Reference Frisch and Hobbs2007) had stated that successful hybridization between different species could be pointers to close evolutionary relationship. However, the low hatchability observed in the hybrids and the percentages of deformed larvae suggest a possible genetic incompatibility between these species from different families. Similar findings have been reported in reciprocal crosses of C. gariepinus with C. batrachus (Sahoo et al., Reference Sahoo, Giri, Sahu and Ayyappan2003; Olufeagba & Okomoda, Reference Olufeagba and Okomoda2016) and in the hybrids between C. gariepinus and Sampa catfish Heterobranchus longifilis Valenciennes, 1840 (Ataguba et al., Reference Ataguba, Annune and Ogbe2009). However, Glamuzina et al. (Reference Glamuzina, Tutman and Skaramuca1999, Reference Glamuzina, Glavić, Skaramuca, Kozul and Tutman2001) observed no difference in the deformity rates of hybrid and non-hybrid groupers, while Owodeinde & Ndimele (Reference Owodeinde and Ndimele2011) reported higher hatchability rates of reciprocal crosses of C. gariepinus and African catfish Heterobranchus bidorsalis Geoffroy Saint-Hilaire, 1809 compared with the pure sibling. The differences observed between these studies could be linked to the differences in the gene compatibility of the species involved in the different crosses reported.
Interspecific hybridization has been thought to be less successful due to the high rate of mortalities during the early life stages (Bartley et al., Reference Bartley, Rena and Immink2001). However, about half the percentage of hatched larvae survived at first feeding. The low survival of the P. hypophthalmus at post-yolk absorption has earlier been justified by the possession of oral teeth and spines, which facilitate grasping of large food items including siblings, hence, accounting for the mortality at this stage (Baras et al., Reference Baras, Slembrouck, Cochet, Caruso and Legendre2010). This is in contrast with the situation in pure Clarias, which never grows long oral teeth, and in which cannibalism essentially takes place at an older age and developmental stage, once sufficient size disparity has taken place (Mollah et al., Reference Mollah, Islam, Faroque and Khan1999). Hence, observations made in this study on cannibalism suggest a preponderant influence of maternal origin upon embryo. This is because mortality linked to cannibalism was more in ♀CG × ♂PH than ♀PH × ♂CG. Many dead fish were found on the bottom of the rearing facility with some missing body parts. This observation is similar to the kind of mortalities described by Solomon & Okomoda (Reference Solomon and Okomoda2012); Appelbaum & McGeer (Reference Appelbaum and McGeer1998) and attributed to cannibalism. However, a recent study by Olufeagba & Okomoda (Reference Olufeagba and Okomoda2016) suggested that hybridization between C. gariepinus and C. batrachus led to significant reduction in cannibalism. Hence, they hypothesized that the genes responsible for cannibalism in C. gariepinus could be recessive to that in C. batrachus. This seems not to be the case in the present study. Nevertheless, in view of the results presented here, larval mortality and thus cannibalism among the hybrids and pure species can be maintained at an ‘acceptably low rate’, if sorting is routinely carried out.
This study shows that reciprocal hybrids can survive up to and reach juvenile stage (and possibly beyond). Nevertheless, it turned out that ♀CG × ♂PH hybrids were more sensitive than others to a degradation of water quality, for reasons that remain to be clearly elucidated. However, the reduced fitness and tolerance of the ♀CG × ♂PH may be linked to a hybridization effect which was earlier reflected in high abnormality percentages in the hatched larvae. Tarnchalanukit (Reference Tarnchalanukit1985) had earlier reported that the hybrid between ♂P. hypophthalmus and ♀C. macrocephalus survived up to 4½ months but was less tolerant to poor water quality. Hence, the issue of survival for the novel hybrid ♀CG × ♂PH might outweigh any other advantage over the parent species whenever water quality is not optimal. The poor water quality observed under the fishmeal-based feeding regime as compared with Artemia or commercial micro-pellets diet justifies the need to optimise feed types administered to larvae of fishes in their early life. The differences in the water stability of the feeds are the likely reasons for the discrepancies observed in the water quality of the system under these feeding regimes.
Table 6 Mean water quality parameters of the experimental unit during the Artemia/micro-pellet diet regime and the fishmeal diet regime

b Means in the same row with different superscript letters differ significantly (P ≤ 0.05).
Many researchers have earlier proposed better performance of reciprocal hybrids over their pure breeds (Madu & Ita, Reference Madu and Ita1991; Jantrarotai, Reference Jantrarotai1993; Tober et al., Reference Tober, Gabriele and Hans-Jurgen1995; Ataguba et al., Reference Ataguba, Annune and Ogbe2010, Solomon et al Reference Solomon, Okomoda and Ochai2013; Olufeagba et al., Reference Olufeagba, Okomoda and Shaibu2016; Olufeagba & Okomoda, Reference Olufeagba and Okomoda2016). However, Chevassus (Reference Chevassus1983) stated that growth of hybrids mostly appears to be intermediate between that of parental species because of partial transmission of traits of the parent to the hybrids. The findings of this study, revealed a positive heterosis for growth in ♀CG × ♂PH and a negative heterosis for ♀PH × ♂CG. The pattern of inheritance as a result of the different crosses may have resulted in the differences in performance observed for the reciprocal crosses. Bartley et al. (Reference Bartley, Rana and Immink1997) had earlier opined that the high preference of Thai fish farmers for hybrids catfishes (between Thai catfish C. macrocephalus and African catfish C. gariepinus) is due to the possession of desirable qualities from both parents. This could be a pointer to possible acceptability of the ♀CG × ♂PH as a large proportion of this hybrid possesses combined features from both parents.
Morphological observations of the hybrids showed two morphotype in the ♀CG × ♂PH hybrid and one in the ♀PH × ♂CG hybrid. The number of morphotypes observed in this study is less compared with the morphotypes reported by Tarnchalanukit (Reference Tarnchalanukit1986), Boonbrahm et al. (Reference Boonbrahm, Tarnchalanukit and Suraniranat1977) and Na-Nakorn et al. (Reference Na-Nakorn and Rangsin1993). However, it is possible that the reduced morphotypes number observed in this study was due to mortalities observed before 35 dph. In line with the morphotype naming format used in similar studies by Tarnchalanukit (Reference Tarnchalanukit1986), Boonbrahm et al. (Reference Boonbrahm, Tarnchalanukit and Suraniranat1977) and Na-Nakorn et al. (Reference Na-Nakorn and Rangsin1993), the Clarias-like hybrids (which is about 13% in the ♀CG × ♂PH and 100% of the ♀PH × ♂CG) were indistinguishable from the pure C. gariepinus. However, the Panga-like hybrid found in the ♀CG × ♂PH showed evidence of shared features (significant modification in fin configuration) from both parents but look like the Pangasius in many ways. Chevassus (Reference Chevassus1983) and Wilkins et al. (Reference Wilkins, Courtney, Gosling, Linnane, Jordan and Curatolo1994) had earlier opined that offspring of interspecific hybridization displayed intermediate morphological characteristics of maternal and paternal species. However, the observation of the two different morphotypes in the ♀CG × ♂PH suggests the possible presence of ploidy polymorphism in the hybrid pool. This is a usually phenomenon associated with many previously reported distant hybridizations trials (Kurita, et al., Reference Kurita, Oshiro, Takashima and Sakaizumi1995; Pandian & Koteeswaran Reference Pandian and Koteeswaran1998; Liu et al., Reference Liu, Liu, You, Chen, Liu, Liu and Liu2010). Furthermore the study by Na-Nakorn et al. (Reference Na-Nakorn and Rangsin1993), on the chromosome characteristics of hybrids between ♂P. hypophthalmus × ♀C. macrocephalus confirmed the presence of diploid, triploid and aneuploidy hybrids corresponding respectively to two intermediate morphotypes (pangasiid-like and clariid-like) and one morphotype indistinguishable from its clariid parent.
Conclusion
The intergeneric cross of Asian catfish P. hypophthalmus and African catfish C. gariepinus in this study produced viable hybrids with potential for culture. However, some hybrids demonstrate lower tolerance to poor water quality when compared with the pure sibs. Future research could be focused on characterising the hybrids and determining the developmental and physiological traits in early and later life stage. Furthermore, the performance of hybrid based on the phenotypic morphotypes can be assessed and compared. However, it is important to state that hybridization between geographically distant species such as this should be conducted very carefully, because escapees from fish farms could just have dramatic and largely unexpected consequences upon wild populations.
Acknowledgements
The authors are indebted to the School of Fisheries and Aquaculture Sciences, Universiti Malaysia Terengganu, Malaysia for providing P. hypophthalmus broodstock used in this study. We also acknowledge the help of Mr. Shaibu Ibrahim, Mr. Giwa-ajeniya Abdullatif and all technical staffs of the PPSPA hatchery department during the breeding trial of this study. This study is part of the first author's Ph.D research.









 Open access
Open access