



1 Introduction
Large organisms propel themselves through a fluid by utilising the inertia of the surrounding fluid. For very small organisms and cells swimming at low Reynolds numbers, inertial propulsion is not possible (Fauci & Dillon Reference Fauci and Dillon2006; Lauga & Powers Reference Lauga and Powers2009); time-reversible kinematics result in no net displacement for the small body. Taylor’s ‘swimming sheet’ is one of the classical models of zero-Reynolds-number swimming; time-reversal symmetry is broken by the wave direction.
Many of the biological fluids in which these cells and organisms swim are non-Newtonian; hence, modelling of swimming in such fluids is of interest. The present study is motivated by the fibrous nature of many biological media, for example, the cervical mucus encountered by the spermatozoa of many internally fertilising species, and active suspensions of elongated cells. Throughout the menstrual cycle, the rheology of cervical mucus changes due to hormonally induced variations in hydration and associated changes in the glycofilament mucin structure. During ovulation, these fibres form a parallel network (figure 1), and sperm migration occurs through this glycofilament structure (Chrétien Reference Chrétien, Hafez and Kenemans1982; Ceric, Silva & Vigil Reference Ceric, Silva and Vigil2005). It is therefore of great interest to determine how Stokesian swimming is modified by the presence of an aligned fibrous network.

Figure 1. Parallel filament mesh in cervical mucus during the time of ovulation;
 $\text{bar}=10~\unicode[STIX]{x03BC}\text{m}$
. Republished with permission of Oxford University Press, from ‘Ultrastructure of the Human Periovulatory Cervical Mucus’, Ceric et al. (Reference Ceric, Silva and Vigil2005); permission conveyed through Copyright Clearance Center, Inc.
$\text{bar}=10~\unicode[STIX]{x03BC}\text{m}$
. Republished with permission of Oxford University Press, from ‘Ultrastructure of the Human Periovulatory Cervical Mucus’, Ceric et al. (Reference Ceric, Silva and Vigil2005); permission conveyed through Copyright Clearance Center, Inc.
Taylor’s pioneering study of Stokesian swimming consists of an infinite sheet undergoing waves of lateral displacement (figure 2). This model was formulated as the far-field Stokes flow produced by a swimming motion given by a small-amplitude sinusoidal wave, and the associated mean rate of working was calculated as a measure of the energetic cost of swimming. Subsequent studies included a 3D model of a waving cylindrical tail (Taylor Reference Taylor1952), investigations by other authors into larger-amplitude motion (Drummond Reference Drummond1966) and more recently the unsteady Stokes flow problem (Pak & Lauga Reference Pak and Lauga2010).
Generalisation of Taylor’s model to non-Newtonian fluids has been an area of significant interest; for a detailed review see Lauga & Powers (Reference Lauga and Powers2009). Chaudhury (Reference Chaudhury1979) initially extended the model to incorporate viscoelastic fluids; it was found that the properties of the fluid lead to an increased steady swimming velocity for lower Reynolds numbers. This problem was reconsidered more recently by Lauga (Reference Lauga2007), who deduced that the mean swimming velocity in a nonlinear viscoelastic fluid is reduced relative to that in a Newtonian fluid; in certain cases the swimming direction is reversed (Fu, Powers & Wolgemuth Reference Fu, Powers and Wolgemuth2007); see also Fu, Wolgemuth & Powers (Reference Fu, Wolgemuth and Powers2009) and Teran, Fauci & Shelley (Reference Teran, Fauci and Shelley2010). Vélez-Cordero & Lauga (Reference Vélez-Cordero and Lauga2013) found propulsion in shear-thinning fluids to be more efficient than that in Newtonian or shear-thickening fluids. Riley & Lauga (Reference Riley and Lauga2014) modelled active propulsion with fluid–structure interaction, and in a subsequent study deduced that for multiple travelling waves, the mean swimming velocity of the sheet is enhanced (Riley & Lauga Reference Riley and Lauga2015). Further to this, Krieger, Spagnolie & Powers (Reference Krieger, Spagnolie and Powers2014), Krieger, Dias & Powers (Reference Krieger, Dias and Powers2015a ) and Krieger, Spagnolie & Powers (Reference Krieger, Spagnolie and Powers2015b ) considered how liquid crystals affect the swimming of micro-organisms. Steady-state and startup models for hexatic liquid crystals were considered along with a nematic steady-state model.
A transversely isotropic fluid exhibits a (perhaps spatially and temporally varying) preferred direction, and has been used to model fibre-reinforced fluids. Previous applications include the mechanical behaviour of collagen gel, the growth of plant root cell walls, suspensions of biomolecules and a multiphase model of extracellular matrix (Green & Friedman Reference Green and Friedman2008; Dyson & Jensen Reference Dyson and Jensen2010; Dyson et al. Reference Dyson, Green, Whiteley and Byrne2015; Holloway, Dyson & Smith Reference Holloway, Dyson and Smith2015). These models comprise a modified constitutive equation describing a viscous fluid with suspended aligned fibres and an expression for the evolution of fibre orientation. A transversely isotropic fluid also provides a model of ‘active’ suspensions of elongated swimmers (Holloway et al. Reference Holloway, Cupples, Smith, Green, Clarke and Dyson2016).

Figure 2. A schematic of Taylor’s swimming sheet in a Newtonian fluid. Working in a frame of reference in which the sheet is stationary,
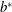 $b^{\ast }$
is the amplitude,
$b^{\ast }$
is the amplitude,
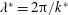 $\unicode[STIX]{x1D706}^{\ast }=2\unicode[STIX]{x03C0}/k^{\ast }$
is the wavelength and
$\unicode[STIX]{x1D706}^{\ast }=2\unicode[STIX]{x03C0}/k^{\ast }$
is the wavelength and
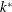 $k^{\ast }$
is the wavenumber. The flow at infinity in the
$k^{\ast }$
is the wavenumber. The flow at infinity in the
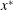 $x^{\ast }$
-direction is
$x^{\ast }$
-direction is
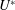 $U^{\ast }$
. A travelling wave traverses the sheet with speed
$U^{\ast }$
. A travelling wave traverses the sheet with speed
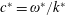 $c^{\ast }=\unicode[STIX]{x1D714}^{\ast }/k^{\ast }$
, where
$c^{\ast }=\unicode[STIX]{x1D714}^{\ast }/k^{\ast }$
, where
 $\unicode[STIX]{x1D714}^{\ast }$
is the angular velocity.
$\unicode[STIX]{x1D714}^{\ast }$
is the angular velocity.
In this study, we consider swimming in transversely isotropic fluids via the constitutive law of Ericksen (Reference Ericksen1960). In § 2, we introduce the governing equations associated with transversely isotropic fluids. The problem is solved in § 3, solving for the leading-order velocity field, mean swimming velocity at next order and mean rate of working. The dependence of these quantities on the rheological parameters is explored in § 4 and discussed in § 5.
2 Governing equations
The incompressibility and generalised Navier–Stokes equations are
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D735}^{\ast }\boldsymbol{\cdot }\boldsymbol{u}^{\ast }=0, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D735}^{\ast }\boldsymbol{\cdot }\boldsymbol{u}^{\ast }=0, & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D70C}^{\ast }\left(\frac{\unicode[STIX]{x2202}\boldsymbol{u}^{\ast }}{\unicode[STIX]{x2202}t^{\ast }}+(\boldsymbol{u}^{\ast }\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast })\boldsymbol{u}^{\ast }\right)=\unicode[STIX]{x1D735}^{\ast }\boldsymbol{\cdot }\unicode[STIX]{x1D748}^{\ast }, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D70C}^{\ast }\left(\frac{\unicode[STIX]{x2202}\boldsymbol{u}^{\ast }}{\unicode[STIX]{x2202}t^{\ast }}+(\boldsymbol{u}^{\ast }\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast })\boldsymbol{u}^{\ast }\right)=\unicode[STIX]{x1D735}^{\ast }\boldsymbol{\cdot }\unicode[STIX]{x1D748}^{\ast }, & \displaystyle\end{eqnarray}$$
where
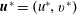 $\boldsymbol{u}^{\ast }=(u^{\ast },v^{\ast })$
is the velocity,
$\boldsymbol{u}^{\ast }=(u^{\ast },v^{\ast })$
is the velocity,
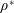 $\unicode[STIX]{x1D70C}^{\ast }$
is the density and
$\unicode[STIX]{x1D70C}^{\ast }$
is the density and
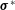 $\unicode[STIX]{x1D748}^{\ast }$
is the stress tensor. We work in a 2D Cartesian coordinate system
$\unicode[STIX]{x1D748}^{\ast }$
is the stress tensor. We work in a 2D Cartesian coordinate system
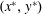 $(x^{\ast },y^{\ast })$
; asterisk notation represents dimensional variables. A constitutive law is also required for
$(x^{\ast },y^{\ast })$
; asterisk notation represents dimensional variables. A constitutive law is also required for
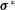 $\unicode[STIX]{x1D748}^{\ast }$
, which we prescribe in the next section.
$\unicode[STIX]{x1D748}^{\ast }$
, which we prescribe in the next section.
2.1 Transversely isotropic stress tensor
Ericksen’s (Reference Ericksen1960) model consists of a stress tensor which is linear in strain rate and depends on a unit vector
 $\boldsymbol{a}$
describing the fibre orientation; this model takes the simplest form that satisfies the required invariances,
$\boldsymbol{a}$
describing the fibre orientation; this model takes the simplest form that satisfies the required invariances,
 $$\begin{eqnarray}\unicode[STIX]{x1D70E}_{ij}^{\ast }=-p^{\ast }\unicode[STIX]{x1D6FF}_{ij}+2\unicode[STIX]{x1D707}^{\ast }e_{ij}^{\ast }+\unicode[STIX]{x1D707}_{1}^{\ast }a_{i}a_{j}+\unicode[STIX]{x1D707}_{2}^{\ast }a_{i}a_{j}a_{k}a_{l}e_{kl}^{\ast }+2\unicode[STIX]{x1D707}_{3}^{\ast }(a_{l}a_{i}e_{lj}^{\ast }+a_{m}a_{j}e_{im}^{\ast }).\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D70E}_{ij}^{\ast }=-p^{\ast }\unicode[STIX]{x1D6FF}_{ij}+2\unicode[STIX]{x1D707}^{\ast }e_{ij}^{\ast }+\unicode[STIX]{x1D707}_{1}^{\ast }a_{i}a_{j}+\unicode[STIX]{x1D707}_{2}^{\ast }a_{i}a_{j}a_{k}a_{l}e_{kl}^{\ast }+2\unicode[STIX]{x1D707}_{3}^{\ast }(a_{l}a_{i}e_{lj}^{\ast }+a_{m}a_{j}e_{im}^{\ast }).\end{eqnarray}$$
We define
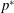 $p^{\ast }$
as the pressure,
$p^{\ast }$
as the pressure,
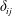 $\unicode[STIX]{x1D6FF}_{ij}$
as the Kronecker delta function and
$\unicode[STIX]{x1D6FF}_{ij}$
as the Kronecker delta function and
 $e_{ij}^{\ast }=(\unicode[STIX]{x2202}u_{i}^{\ast }/\unicode[STIX]{x2202}x_{j}^{\ast }+\unicode[STIX]{x2202}u_{j}^{\ast }/\unicode[STIX]{x2202}x_{i}^{\ast })/2$
as the rate-of-strain tensor (Ericksen Reference Ericksen1960).
$e_{ij}^{\ast }=(\unicode[STIX]{x2202}u_{i}^{\ast }/\unicode[STIX]{x2202}x_{j}^{\ast }+\unicode[STIX]{x2202}u_{j}^{\ast }/\unicode[STIX]{x2202}x_{i}^{\ast })/2$
as the rate-of-strain tensor (Ericksen Reference Ericksen1960).
By considering simple flows with a uniform director field, we may interpret the rheological parameters as follows. By setting
 $\unicode[STIX]{x1D707}_{1}^{\ast }=\unicode[STIX]{x1D707}_{2}^{\ast }=\unicode[STIX]{x1D707}_{3}^{\ast }=0$
, the stress tensor for an incompressible Newtonian fluid remains, with ‘matrix viscosity’
$\unicode[STIX]{x1D707}_{1}^{\ast }=\unicode[STIX]{x1D707}_{2}^{\ast }=\unicode[STIX]{x1D707}_{3}^{\ast }=0$
, the stress tensor for an incompressible Newtonian fluid remains, with ‘matrix viscosity’
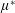 $\unicode[STIX]{x1D707}^{\ast }$
(Holloway et al.
Reference Holloway, Dyson and Smith2015). The term with
$\unicode[STIX]{x1D707}^{\ast }$
(Holloway et al.
Reference Holloway, Dyson and Smith2015). The term with
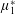 $\unicode[STIX]{x1D707}_{1}^{\ast }$
has no dependence on velocity, suggesting that
$\unicode[STIX]{x1D707}_{1}^{\ast }$
has no dependence on velocity, suggesting that
 $\unicode[STIX]{x1D707}_{1}^{\ast }$
relates to a tension in the fibre direction (Dyson & Jensen Reference Dyson and Jensen2010). This term can also be related to the stresslet-type active behaviour of fibres in a perfectly aligned active fluid (Holloway et al.
Reference Holloway, Cupples, Smith, Green, Clarke and Dyson2016). We will therefore refer to this quantity as the active parameter. This term can be taken as a simple model for suspensions of self-propelling microscopic bodies such as bacteria or active gels of molecular motor proteins. The viscosity associated with extensional flow parallel to the fibre direction is
$\unicode[STIX]{x1D707}_{1}^{\ast }$
relates to a tension in the fibre direction (Dyson & Jensen Reference Dyson and Jensen2010). This term can also be related to the stresslet-type active behaviour of fibres in a perfectly aligned active fluid (Holloway et al.
Reference Holloway, Cupples, Smith, Green, Clarke and Dyson2016). We will therefore refer to this quantity as the active parameter. This term can be taken as a simple model for suspensions of self-propelling microscopic bodies such as bacteria or active gels of molecular motor proteins. The viscosity associated with extensional flow parallel to the fibre direction is
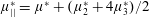 $\unicode[STIX]{x1D707}_{||}^{\ast }=\unicode[STIX]{x1D707}^{\ast }+(\unicode[STIX]{x1D707}_{2}^{\ast }+4\unicode[STIX]{x1D707}_{3}^{\ast })/2$
, the viscosity associated with the flow orthogonal to the fibre direction is
$\unicode[STIX]{x1D707}_{||}^{\ast }=\unicode[STIX]{x1D707}^{\ast }+(\unicode[STIX]{x1D707}_{2}^{\ast }+4\unicode[STIX]{x1D707}_{3}^{\ast })/2$
, the viscosity associated with the flow orthogonal to the fibre direction is
 $\unicode[STIX]{x1D707}_{\bot }^{\ast }=\unicode[STIX]{x1D707}^{\ast }$
and the viscosity of shear flow in the fibre direction is
$\unicode[STIX]{x1D707}_{\bot }^{\ast }=\unicode[STIX]{x1D707}^{\ast }$
and the viscosity of shear flow in the fibre direction is
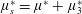 $\unicode[STIX]{x1D707}_{s}^{\ast }=\unicode[STIX]{x1D707}^{\ast }+\unicode[STIX]{x1D707}_{3}^{\ast }$
(Dyson & Jensen Reference Dyson and Jensen2010). Since
$\unicode[STIX]{x1D707}_{s}^{\ast }=\unicode[STIX]{x1D707}^{\ast }+\unicode[STIX]{x1D707}_{3}^{\ast }$
(Dyson & Jensen Reference Dyson and Jensen2010). Since
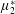 $\unicode[STIX]{x1D707}_{2}^{\ast }$
only has an impact on extensional viscosity parallel to the fibre direction,
$\unicode[STIX]{x1D707}_{2}^{\ast }$
only has an impact on extensional viscosity parallel to the fibre direction,
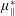 $\unicode[STIX]{x1D707}_{\Vert }^{\ast }$
, it is termed the anisotropic extensional viscosity. The parameter
$\unicode[STIX]{x1D707}_{\Vert }^{\ast }$
, it is termed the anisotropic extensional viscosity. The parameter
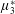 $\unicode[STIX]{x1D707}_{3}^{\ast }$
distinguishes
$\unicode[STIX]{x1D707}_{3}^{\ast }$
distinguishes
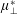 $\unicode[STIX]{x1D707}_{\bot }^{\ast }$
from
$\unicode[STIX]{x1D707}_{\bot }^{\ast }$
from
 $\unicode[STIX]{x1D707}_{s}^{\ast }$
and so is labelled the anisotropic shear viscosity; this parameter represents the difference between shear viscosities parallel and perpendicular to the fibre direction (Green & Friedman Reference Green and Friedman2008; Dyson & Jensen Reference Dyson and Jensen2010; Holloway et al.
Reference Holloway, Dyson and Smith2015).
$\unicode[STIX]{x1D707}_{s}^{\ast }$
and so is labelled the anisotropic shear viscosity; this parameter represents the difference between shear viscosities parallel and perpendicular to the fibre direction (Green & Friedman Reference Green and Friedman2008; Dyson & Jensen Reference Dyson and Jensen2010; Holloway et al.
Reference Holloway, Dyson and Smith2015).
2.2 Fibre evolution equation
A fibre evolution equation describes the evolution of fibre orientation with time. We use the form given by Green & Friedman (Reference Green and Friedman2008),
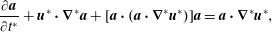 $$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\boldsymbol{a}}{\unicode[STIX]{x2202}t^{\ast }}+\boldsymbol{u}^{\ast }\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{a}+[\boldsymbol{a}\boldsymbol{\cdot }(\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{u}^{\ast })]\boldsymbol{a}=\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{u}^{\ast },\end{eqnarray}$$
$$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\boldsymbol{a}}{\unicode[STIX]{x2202}t^{\ast }}+\boldsymbol{u}^{\ast }\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{a}+[\boldsymbol{a}\boldsymbol{\cdot }(\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{u}^{\ast })]\boldsymbol{a}=\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{u}^{\ast },\end{eqnarray}$$
which corresponds to a specific case of Ericksen’s (Reference Ericksen1960) equation in the long-fibre limit. It should be noted that
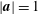 $|\boldsymbol{a}|=1$
, and thus the model only considers local alignment of fibres and not their length. This gives a generalised form of Jeffery’s treatment for long ellipsoidal particles aligning with flow (Jeffery Reference Jeffery1922; Dyson & Jensen Reference Dyson and Jensen2010).
$|\boldsymbol{a}|=1$
, and thus the model only considers local alignment of fibres and not their length. This gives a generalised form of Jeffery’s treatment for long ellipsoidal particles aligning with flow (Jeffery Reference Jeffery1922; Dyson & Jensen Reference Dyson and Jensen2010).
Since
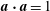 $\boldsymbol{a}\boldsymbol{\cdot }\boldsymbol{a}=1$
, the component of (2.4) in the
$\boldsymbol{a}\boldsymbol{\cdot }\boldsymbol{a}=1$
, the component of (2.4) in the
 $\boldsymbol{a}$
-direction is automatically satisfied. The orthogonal component of (2.4) is
$\boldsymbol{a}$
-direction is automatically satisfied. The orthogonal component of (2.4) is
 $$\begin{eqnarray}\boldsymbol{a}^{\bot }\boldsymbol{\cdot }\left[\frac{\unicode[STIX]{x2202}\boldsymbol{a}}{\unicode[STIX]{x2202}t^{\ast }}+\boldsymbol{u}^{\ast }\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{a}+[\boldsymbol{a}\boldsymbol{\cdot }(\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{u}^{\ast })]\boldsymbol{a}-\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{u}^{\ast }\right]=0,\end{eqnarray}$$
$$\begin{eqnarray}\boldsymbol{a}^{\bot }\boldsymbol{\cdot }\left[\frac{\unicode[STIX]{x2202}\boldsymbol{a}}{\unicode[STIX]{x2202}t^{\ast }}+\boldsymbol{u}^{\ast }\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{a}+[\boldsymbol{a}\boldsymbol{\cdot }(\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{u}^{\ast })]\boldsymbol{a}-\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}^{\ast }\boldsymbol{u}^{\ast }\right]=0,\end{eqnarray}$$
where
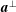 $\boldsymbol{a}^{\bot }$
is a unit vector perpendicular to
$\boldsymbol{a}^{\bot }$
is a unit vector perpendicular to
 $\boldsymbol{a}$
.
$\boldsymbol{a}$
.
2.3 Boundary conditions
We work in a frame of reference moving with the swimmer in the
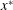 $x^{\ast }$
-direction; the horizontal flow as
$x^{\ast }$
-direction; the horizontal flow as
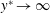 $y^{\ast }\rightarrow \infty$
therefore gives the mean swimming velocity. No-slip conditions on the sheet, representing a travelling wave with speed
$y^{\ast }\rightarrow \infty$
therefore gives the mean swimming velocity. No-slip conditions on the sheet, representing a travelling wave with speed
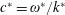 $c^{\ast }=\unicode[STIX]{x1D714}^{\ast }/k^{\ast }$
, are thus
$c^{\ast }=\unicode[STIX]{x1D714}^{\ast }/k^{\ast }$
, are thus
 $$\begin{eqnarray}u^{\ast }=0,\quad v^{\ast }=-\unicode[STIX]{x1D714}^{\ast }b^{\ast }\cos (k^{\ast }x^{\ast }-\unicode[STIX]{x1D714}^{\ast }t^{\ast }),\quad \text{on }y^{\ast }=y_{s}^{\ast }=b^{\ast }\sin (k^{\ast }x^{\ast }-\unicode[STIX]{x1D714}^{\ast }t^{\ast }).\end{eqnarray}$$
$$\begin{eqnarray}u^{\ast }=0,\quad v^{\ast }=-\unicode[STIX]{x1D714}^{\ast }b^{\ast }\cos (k^{\ast }x^{\ast }-\unicode[STIX]{x1D714}^{\ast }t^{\ast }),\quad \text{on }y^{\ast }=y_{s}^{\ast }=b^{\ast }\sin (k^{\ast }x^{\ast }-\unicode[STIX]{x1D714}^{\ast }t^{\ast }).\end{eqnarray}$$
The parameter
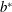 $b^{\ast }$
is the amplitude,
$b^{\ast }$
is the amplitude,
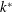 $k^{\ast }$
is the wavenumber,
$k^{\ast }$
is the wavenumber,
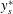 $y_{s}^{\ast }$
is the equation of the sheet surface and
$y_{s}^{\ast }$
is the equation of the sheet surface and
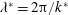 $\unicode[STIX]{x1D706}^{\ast }=2\unicode[STIX]{x03C0}/k^{\ast }$
is the wavelength. The velocity must remain bounded as
$\unicode[STIX]{x1D706}^{\ast }=2\unicode[STIX]{x03C0}/k^{\ast }$
is the wavelength. The velocity must remain bounded as
 $y^{\ast }\rightarrow \infty$
.
$y^{\ast }\rightarrow \infty$
.
2.4 Non-dimensionalisation
The model is non-dimensionalised as follows:
 $$\begin{eqnarray}\boldsymbol{u}^{\ast }=\frac{\unicode[STIX]{x1D714}^{\ast }}{k^{\ast }}\boldsymbol{u},\quad \boldsymbol{x}^{\ast }=\frac{\boldsymbol{x}}{k^{\ast }},\quad t^{\ast }=\frac{t}{\unicode[STIX]{x1D714}^{\ast }},\quad p^{\ast }=\unicode[STIX]{x1D714}^{\ast }\unicode[STIX]{x1D707}^{\ast }p,\quad \unicode[STIX]{x1D748}^{\ast }=\unicode[STIX]{x1D714}^{\ast }\unicode[STIX]{x1D707}^{\ast }\unicode[STIX]{x1D748}.\end{eqnarray}$$
$$\begin{eqnarray}\boldsymbol{u}^{\ast }=\frac{\unicode[STIX]{x1D714}^{\ast }}{k^{\ast }}\boldsymbol{u},\quad \boldsymbol{x}^{\ast }=\frac{\boldsymbol{x}}{k^{\ast }},\quad t^{\ast }=\frac{t}{\unicode[STIX]{x1D714}^{\ast }},\quad p^{\ast }=\unicode[STIX]{x1D714}^{\ast }\unicode[STIX]{x1D707}^{\ast }p,\quad \unicode[STIX]{x1D748}^{\ast }=\unicode[STIX]{x1D714}^{\ast }\unicode[STIX]{x1D707}^{\ast }\unicode[STIX]{x1D748}.\end{eqnarray}$$
The continuity and fibre evolution equations are unchanged. For microscopic swimmers, the Reynolds number,
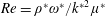 $Re=\unicode[STIX]{x1D70C}^{\ast }\unicode[STIX]{x1D714}^{\ast }/{k^{\ast }}^{2}\unicode[STIX]{x1D707}^{\ast }$
, is much less than 1, so we neglect inertial terms. The resulting system of partial differential equations is therefore
$Re=\unicode[STIX]{x1D70C}^{\ast }\unicode[STIX]{x1D714}^{\ast }/{k^{\ast }}^{2}\unicode[STIX]{x1D707}^{\ast }$
, is much less than 1, so we neglect inertial terms. The resulting system of partial differential equations is therefore
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D735}\boldsymbol{\cdot }\boldsymbol{u}=0, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D735}\boldsymbol{\cdot }\boldsymbol{u}=0, & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D735}\boldsymbol{\cdot }\unicode[STIX]{x1D748}=\mathbf{0}, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D735}\boldsymbol{\cdot }\unicode[STIX]{x1D748}=\mathbf{0}, & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle \boldsymbol{a}^{\bot }\boldsymbol{\cdot }\left[\frac{\unicode[STIX]{x2202}\boldsymbol{a}}{\unicode[STIX]{x2202}t}+\boldsymbol{u}\boldsymbol{\cdot }\unicode[STIX]{x1D735}\boldsymbol{a}+[\boldsymbol{a}\boldsymbol{\cdot }(\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}\boldsymbol{u})]\boldsymbol{a}-\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}\boldsymbol{u}\right]=0, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \boldsymbol{a}^{\bot }\boldsymbol{\cdot }\left[\frac{\unicode[STIX]{x2202}\boldsymbol{a}}{\unicode[STIX]{x2202}t}+\boldsymbol{u}\boldsymbol{\cdot }\unicode[STIX]{x1D735}\boldsymbol{a}+[\boldsymbol{a}\boldsymbol{\cdot }(\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}\boldsymbol{u})]\boldsymbol{a}-\boldsymbol{a}\boldsymbol{\cdot }\unicode[STIX]{x1D735}\boldsymbol{u}\right]=0, & \displaystyle\end{eqnarray}$$
where
 $$\begin{eqnarray}\unicode[STIX]{x1D70E}_{ij}=-p\unicode[STIX]{x1D6FF}_{ij}+2e_{ij}+\unicode[STIX]{x1D707}_{1}a_{i}a_{j}+\unicode[STIX]{x1D707}_{2}a_{i}a_{j}a_{k}a_{l}e_{kl}+2\unicode[STIX]{x1D707}_{3}(a_{l}a_{i}e_{lj}+a_{m}a_{j}e_{im}),\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D70E}_{ij}=-p\unicode[STIX]{x1D6FF}_{ij}+2e_{ij}+\unicode[STIX]{x1D707}_{1}a_{i}a_{j}+\unicode[STIX]{x1D707}_{2}a_{i}a_{j}a_{k}a_{l}e_{kl}+2\unicode[STIX]{x1D707}_{3}(a_{l}a_{i}e_{lj}+a_{m}a_{j}e_{im}),\end{eqnarray}$$
with dimensionless groups
 $$\begin{eqnarray}\unicode[STIX]{x1D707}_{1}=\frac{\unicode[STIX]{x1D707}_{1}^{\ast }}{\unicode[STIX]{x1D707}^{\ast }\unicode[STIX]{x1D714}^{\ast }},\quad \unicode[STIX]{x1D707}_{2}=\frac{\unicode[STIX]{x1D707}_{2}^{\ast }}{\unicode[STIX]{x1D707}^{\ast }},\quad \unicode[STIX]{x1D707}_{3}=\frac{\unicode[STIX]{x1D707}_{3}^{\ast }}{\unicode[STIX]{x1D707}^{\ast }}.\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D707}_{1}=\frac{\unicode[STIX]{x1D707}_{1}^{\ast }}{\unicode[STIX]{x1D707}^{\ast }\unicode[STIX]{x1D714}^{\ast }},\quad \unicode[STIX]{x1D707}_{2}=\frac{\unicode[STIX]{x1D707}_{2}^{\ast }}{\unicode[STIX]{x1D707}^{\ast }},\quad \unicode[STIX]{x1D707}_{3}=\frac{\unicode[STIX]{x1D707}_{3}^{\ast }}{\unicode[STIX]{x1D707}^{\ast }}.\end{eqnarray}$$
The boundary conditions (2.6) become
 $$\begin{eqnarray}u=0,\quad v=-\unicode[STIX]{x1D700}\cos (x-t),\quad \text{on }y=y_{s}=\unicode[STIX]{x1D700}\sin (x-t),\end{eqnarray}$$
$$\begin{eqnarray}u=0,\quad v=-\unicode[STIX]{x1D700}\cos (x-t),\quad \text{on }y=y_{s}=\unicode[STIX]{x1D700}\sin (x-t),\end{eqnarray}$$
where
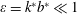 $\unicode[STIX]{x1D700}=k^{\ast }b^{\ast }\ll 1$
. Moreover,
$\unicode[STIX]{x1D700}=k^{\ast }b^{\ast }\ll 1$
. Moreover,
 $u$
and
$u$
and
 $v$
must remain bounded as
$v$
must remain bounded as
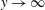 $y\rightarrow \infty$
.
$y\rightarrow \infty$
.

Figure 3. Regimes of interest in the parameter space: (i) the plane
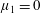 $\unicode[STIX]{x1D707}_{1}=0$
is the passive transversely isotropic regime; (ii) the dashed line represents the active-only regime where
$\unicode[STIX]{x1D707}_{1}=0$
is the passive transversely isotropic regime; (ii) the dashed line represents the active-only regime where
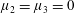 $\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
; (iii) the quarter cylinder is the nearly isotropic regime where neither
$\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
; (iii) the quarter cylinder is the nearly isotropic regime where neither
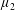 $\unicode[STIX]{x1D707}_{2}$
nor
$\unicode[STIX]{x1D707}_{2}$
nor
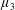 $\unicode[STIX]{x1D707}_{3}$
is large; (iv) the remaining region is the fully active and transversely isotropic regime.
$\unicode[STIX]{x1D707}_{3}$
is large; (iv) the remaining region is the fully active and transversely isotropic regime.
Four regimes in parameter space, depicted in figure 3, will be considered in our results: (i) a passive transversely isotropic fluid, occurring when
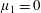 $\unicode[STIX]{x1D707}_{1}=0$
; (ii) an active fluid (
$\unicode[STIX]{x1D707}_{1}=0$
; (ii) an active fluid (
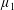 $\unicode[STIX]{x1D707}_{1}$
non-zero) with
$\unicode[STIX]{x1D707}_{1}$
non-zero) with
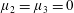 $\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
; (iii) a nearly isotropic regime, where all parameters take values up to
$\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
; (iii) a nearly isotropic regime, where all parameters take values up to
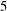 $5$
; (iv) the regime where at least one of
$5$
; (iv) the regime where at least one of
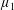 $\unicode[STIX]{x1D707}_{1}$
,
$\unicode[STIX]{x1D707}_{1}$
,
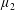 $\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{2}$
and
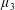 $\unicode[STIX]{x1D707}_{3}$
is much larger than 1. It should be noted that
$\unicode[STIX]{x1D707}_{3}$
is much larger than 1. It should be noted that
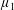 $\unicode[STIX]{x1D707}_{1}$
may be positive or negative, representing active ‘puller’ or ‘pusher’ behaviour respectively (Saintillan Reference Saintillan2010).
$\unicode[STIX]{x1D707}_{1}$
may be positive or negative, representing active ‘puller’ or ‘pusher’ behaviour respectively (Saintillan Reference Saintillan2010).
3 Asymptotic solution

Figure 4. A schematic showing the initial uniform orientation angle,
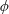 $\unicode[STIX]{x1D719}$
, and the small perturbation away from this angle,
$\unicode[STIX]{x1D719}$
, and the small perturbation away from this angle,
 $\unicode[STIX]{x1D703}$
.
$\unicode[STIX]{x1D703}$
.
3.1 Stream function formulation
To determine the effect of fibres on the mean swimming velocity, we consider an initially spatially uniform fibre angle,
 $\unicode[STIX]{x1D719}$
, aligned such that
$\unicode[STIX]{x1D719}$
, aligned such that
 $\boldsymbol{a}(x,y,0)=(\cos \unicode[STIX]{x1D719},\sin \unicode[STIX]{x1D719})$
. As the sheet swims, this alignment will evolve, initially as a small perturbation
$\boldsymbol{a}(x,y,0)=(\cos \unicode[STIX]{x1D719},\sin \unicode[STIX]{x1D719})$
. As the sheet swims, this alignment will evolve, initially as a small perturbation
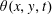 $\unicode[STIX]{x1D703}(x,y,t)$
such that
$\unicode[STIX]{x1D703}(x,y,t)$
such that
 $$\begin{eqnarray}\displaystyle \boldsymbol{a} & = & \displaystyle (\cos (\unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}),\sin (\unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}))\nonumber\\ \displaystyle & {\approx} & \displaystyle (\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719},\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \boldsymbol{a} & = & \displaystyle (\cos (\unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}),\sin (\unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}))\nonumber\\ \displaystyle & {\approx} & \displaystyle (\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719},\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\end{eqnarray}$$
(see figure 4). The components of the stress tensor in terms of
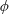 $\unicode[STIX]{x1D719}$
and
$\unicode[STIX]{x1D719}$
and
 $\unicode[STIX]{x1D703}$
are given in appendix A. Taking the curl of (2.9) eliminates pressure, reducing (2.9) to a single equation. Since the flow is incompressible, we introduce a dimensionless stream function
$\unicode[STIX]{x1D703}$
are given in appendix A. Taking the curl of (2.9) eliminates pressure, reducing (2.9) to a single equation. Since the flow is incompressible, we introduce a dimensionless stream function
 $$\begin{eqnarray}u={\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{}}},\quad v=-{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}}},\end{eqnarray}$$
$$\begin{eqnarray}u={\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{}}},\quad v=-{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}}},\end{eqnarray}$$
transforming (2.9) to
 $$\begin{eqnarray}\displaystyle & & \displaystyle \!\!\!\!\left(1+\frac{\unicode[STIX]{x1D707}_{2}}{4}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\right)\unicode[STIX]{x1D6FB}^{4}\unicode[STIX]{x1D713}-\unicode[STIX]{x1D707}_{1}\left[2\sin 2\unicode[STIX]{x1D719}\left(\unicode[STIX]{x1D703}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{2}}}\right)+\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}\right)^{2}-\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}\right)^{2}\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.\vphantom{\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}\right)^{2}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right)+\cos 2\unicode[STIX]{x1D719}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{2}}}+4\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}+\unicode[STIX]{x1D703}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right)\right)\right]\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad -\,\unicode[STIX]{x1D707}_{2}\left[\sin 4\unicode[STIX]{x1D719}\left(-\frac{\unicode[STIX]{x1D703}}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{4}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{4}}}\right)+\frac{1}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{2}}}\right)\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{3}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{}}}\right)+2{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}\left(3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{3}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.+\,\unicode[STIX]{x1D703}{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+\frac{1}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}\right)\right)+\cos 4\unicode[STIX]{x1D719}\left(2\unicode[STIX]{x1D703}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.+\,\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{2}}}\right){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{3}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}\left(3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{3}}}\right)-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\right)-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}\right)\right]=0.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & & \displaystyle \!\!\!\!\left(1+\frac{\unicode[STIX]{x1D707}_{2}}{4}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\right)\unicode[STIX]{x1D6FB}^{4}\unicode[STIX]{x1D713}-\unicode[STIX]{x1D707}_{1}\left[2\sin 2\unicode[STIX]{x1D719}\left(\unicode[STIX]{x1D703}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{2}}}\right)+\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}\right)^{2}-\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}\right)^{2}\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.\vphantom{\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}\right)^{2}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right)+\cos 2\unicode[STIX]{x1D719}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{2}}}+4\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}+\unicode[STIX]{x1D703}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right)\right)\right]\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad -\,\unicode[STIX]{x1D707}_{2}\left[\sin 4\unicode[STIX]{x1D719}\left(-\frac{\unicode[STIX]{x1D703}}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{4}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{4}}}\right)+\frac{1}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{2}}}\right)\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{3}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{}}}\right)+2{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}\left(3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{3}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.+\,\unicode[STIX]{x1D703}{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+\frac{1}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}\right)\right)+\cos 4\unicode[STIX]{x1D719}\left(2\unicode[STIX]{x1D703}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.+\,\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{2}}}\right){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{3}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \!\!\!\!\quad \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}\left(3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{3}}}\right)-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\right)-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}\right)\right]=0.\end{eqnarray}$$
When
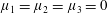 $\unicode[STIX]{x1D707}_{1}=\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
, (3.3) reduces to the familiar biharmonic equation of Newtonian Stokes flow. The evolution equation (2.10) becomes
$\unicode[STIX]{x1D707}_{1}=\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
, (3.3) reduces to the familiar biharmonic equation of Newtonian Stokes flow. The evolution equation (2.10) becomes
 $$\begin{eqnarray}\displaystyle & & \displaystyle {\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}t^{}}}+\left[{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}\right]+\left(\sin ^{2}\unicode[STIX]{x1D719}+\frac{\unicode[STIX]{x1D703}}{2}\sin 2\unicode[STIX]{x1D719}\right){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}\nonumber\\ \displaystyle & & \displaystyle \quad +\,(\sin 2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos 2\unicode[STIX]{x1D719}){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+\left(\cos ^{2}\unicode[STIX]{x1D719}-\frac{\unicode[STIX]{x1D703}}{2}\sin 2\unicode[STIX]{x1D719}\right){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\nonumber\\ \displaystyle & & \displaystyle \quad +\,\unicode[STIX]{x1D703}\left(\left[(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}-(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}\right]{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right.\nonumber\\ \displaystyle & & \displaystyle \left.\quad +\,(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\right)\right)=0.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & & \displaystyle {\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}t^{}}}+\left[{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}x^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}}{\unicode[STIX]{x2202}y^{}}}\right]+\left(\sin ^{2}\unicode[STIX]{x1D719}+\frac{\unicode[STIX]{x1D703}}{2}\sin 2\unicode[STIX]{x1D719}\right){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}\nonumber\\ \displaystyle & & \displaystyle \quad +\,(\sin 2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos 2\unicode[STIX]{x1D719}){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+\left(\cos ^{2}\unicode[STIX]{x1D719}-\frac{\unicode[STIX]{x1D703}}{2}\sin 2\unicode[STIX]{x1D719}\right){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\nonumber\\ \displaystyle & & \displaystyle \quad +\,\unicode[STIX]{x1D703}\left(\left[(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}-(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}\right]{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right.\nonumber\\ \displaystyle & & \displaystyle \left.\quad +\,(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\right)\right)=0.\end{eqnarray}$$
The boundary conditions (2.13) become
 $$\begin{eqnarray}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{}}}=0,\quad {\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}}}=\unicode[STIX]{x1D700}\cos (x-t),\quad \text{on}\,y=\unicode[STIX]{x1D700}\sin (x-t),\end{eqnarray}$$
$$\begin{eqnarray}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{}}}=0,\quad {\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{}}}=\unicode[STIX]{x1D700}\cos (x-t),\quad \text{on}\,y=\unicode[STIX]{x1D700}\sin (x-t),\end{eqnarray}$$
with
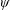 $\unicode[STIX]{x1D713}$
having bounded first derivatives as
$\unicode[STIX]{x1D713}$
having bounded first derivatives as
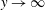 $y\rightarrow \infty$
.
$y\rightarrow \infty$
.
3.2 Perturbation expansion
To apply the boundary conditions at
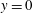 $y=0$
rather than on the sheet, we make the small-amplitude expansion
$y=0$
rather than on the sheet, we make the small-amplitude expansion
 $$\begin{eqnarray}\displaystyle & \displaystyle \left.\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y}\right|_{y=0}+\left.\unicode[STIX]{x1D700}\sin (x-t)\frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}\right|_{y=0}+\cdots =0, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \left.\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y}\right|_{y=0}+\left.\unicode[STIX]{x1D700}\sin (x-t)\frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}\right|_{y=0}+\cdots =0, & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle \left.\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x}\right|_{y=0}+\left.\unicode[STIX]{x1D700}\sin (x-t)\frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y\unicode[STIX]{x2202}x}\right|_{y=0}+\cdots =\unicode[STIX]{x1D700}\cos (x-t). & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \left.\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x}\right|_{y=0}+\left.\unicode[STIX]{x1D700}\sin (x-t)\frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y\unicode[STIX]{x2202}x}\right|_{y=0}+\cdots =\unicode[STIX]{x1D700}\cos (x-t). & \displaystyle\end{eqnarray}$$
The velocity and fibre angle perturbations thus take the form
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D713}(x,y,t;\unicode[STIX]{x1D700})=\unicode[STIX]{x1D700}\unicode[STIX]{x1D713}_{0}(x,y,t)+\unicode[STIX]{x1D700}^{2}\unicode[STIX]{x1D713}_{1}(x,y,t)+\cdots \,, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D713}(x,y,t;\unicode[STIX]{x1D700})=\unicode[STIX]{x1D700}\unicode[STIX]{x1D713}_{0}(x,y,t)+\unicode[STIX]{x1D700}^{2}\unicode[STIX]{x1D713}_{1}(x,y,t)+\cdots \,, & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D703}(x,y,t;\unicode[STIX]{x1D700})=\unicode[STIX]{x1D700}\unicode[STIX]{x1D703}_{0}(x,y,t)+\unicode[STIX]{x1D700}^{2}\unicode[STIX]{x1D703}_{1}(x,y,t)+\cdots \,. & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D703}(x,y,t;\unicode[STIX]{x1D700})=\unicode[STIX]{x1D700}\unicode[STIX]{x1D703}_{0}(x,y,t)+\unicode[STIX]{x1D700}^{2}\unicode[STIX]{x1D703}_{1}(x,y,t)+\cdots \,. & \displaystyle\end{eqnarray}$$
As in Taylor’s analysis, the background flow (i.e. the sheet swimming velocity) will occur at order
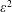 $\unicode[STIX]{x1D700}^{2}$
.
$\unicode[STIX]{x1D700}^{2}$
.
3.3 Leading-order solution
Substituting the expansions into (3.3) and (3.4) and equating coefficients of powers of
 $\unicode[STIX]{x1D700}$
yields the leading-order partial differential equation. At order
$\unicode[STIX]{x1D700}$
yields the leading-order partial differential equation. At order
 $\unicode[STIX]{x1D700}$
, (3.3) yields
$\unicode[STIX]{x1D700}$
, (3.3) yields
 $$\begin{eqnarray}\displaystyle & & \displaystyle \left(1+\frac{\unicode[STIX]{x1D707}_{2}}{4}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\right)\unicode[STIX]{x1D6FB}^{4}\unicode[STIX]{x1D713}_{0}-\unicode[STIX]{x1D707}_{1}\left(2\sin 2\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+\cos 2\unicode[STIX]{x1D719}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{2}}}\right)\right)\nonumber\\ \displaystyle & & \displaystyle \quad +\,\unicode[STIX]{x1D707}_{2}\left(\cos 4\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+\frac{\sin 4\unicode[STIX]{x1D719}}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}\right)\right)=0,\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & & \displaystyle \left(1+\frac{\unicode[STIX]{x1D707}_{2}}{4}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\right)\unicode[STIX]{x1D6FB}^{4}\unicode[STIX]{x1D713}_{0}-\unicode[STIX]{x1D707}_{1}\left(2\sin 2\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+\cos 2\unicode[STIX]{x1D719}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{2}}}\right)\right)\nonumber\\ \displaystyle & & \displaystyle \quad +\,\unicode[STIX]{x1D707}_{2}\left(\cos 4\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+\frac{\sin 4\unicode[STIX]{x1D719}}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}\right)\right)=0,\end{eqnarray}$$
and (3.4) yields
 $$\begin{eqnarray}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}t^{}}}+\sin 2\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+\cos ^{2}\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}}}+\sin ^{2}\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{2}}}=0.\end{eqnarray}$$
$$\begin{eqnarray}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}t^{}}}+\sin 2\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+\cos ^{2}\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}}}+\sin ^{2}\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{2}}}=0.\end{eqnarray}$$
The boundary conditions (3.6) and (3.7) become
 $$\begin{eqnarray}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{}}}=0,\quad {\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}}}=\cos (x-t),\quad \text{on }y=0,\end{eqnarray}$$
$$\begin{eqnarray}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{}}}=0,\quad {\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}}}=\cos (x-t),\quad \text{on }y=0,\end{eqnarray}$$
combined with the requirement that the derivatives of
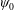 $\unicode[STIX]{x1D713}_{0}$
are bounded as
$\unicode[STIX]{x1D713}_{0}$
are bounded as
 $y\rightarrow \infty$
.
$y\rightarrow \infty$
.
Equations (3.10) and (3.11) are solved with the ansatz
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D713}_{0}=f_{1}(y)\cos (x-t)+f_{2}(y)\sin (x-t), & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D713}_{0}=f_{1}(y)\cos (x-t)+f_{2}(y)\sin (x-t), & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D703}_{0}=g_{1}(y)\cos (x-t)+g_{2}(y)\sin (x-t), & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D703}_{0}=g_{1}(y)\cos (x-t)+g_{2}(y)\sin (x-t), & \displaystyle\end{eqnarray}$$
for some functions
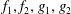 $f_{1},f_{2},g_{1},g_{2}$
. Comparing coefficients of sine and cosine leads to a system of four ordinary differential equations,
$f_{1},f_{2},g_{1},g_{2}$
. Comparing coefficients of sine and cosine leads to a system of four ordinary differential equations,
 $$\begin{eqnarray}\displaystyle & & \displaystyle (1+{\textstyle \frac{1}{4}}\unicode[STIX]{x1D707}_{2}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3})(f_{1}^{\prime \prime \prime \prime }-2f_{1}^{\prime \prime }+f_{1})+\unicode[STIX]{x1D707}_{1}(\cos 2\unicode[STIX]{x1D719}\,(g_{1}+g_{1}^{\prime \prime })-2\sin 2\unicode[STIX]{x1D719}\,g_{1}^{\prime })\nonumber\\ \displaystyle & & \displaystyle \quad +\,\unicode[STIX]{x1D707}_{2}({\textstyle \frac{1}{2}}\sin 4\unicode[STIX]{x1D719}\,(f_{2}^{\prime \prime \prime }+f_{2}^{\prime \prime })-\cos 4\unicode[STIX]{x1D719}\,f_{1}^{\prime \prime })=0,\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & & \displaystyle (1+{\textstyle \frac{1}{4}}\unicode[STIX]{x1D707}_{2}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3})(f_{1}^{\prime \prime \prime \prime }-2f_{1}^{\prime \prime }+f_{1})+\unicode[STIX]{x1D707}_{1}(\cos 2\unicode[STIX]{x1D719}\,(g_{1}+g_{1}^{\prime \prime })-2\sin 2\unicode[STIX]{x1D719}\,g_{1}^{\prime })\nonumber\\ \displaystyle & & \displaystyle \quad +\,\unicode[STIX]{x1D707}_{2}({\textstyle \frac{1}{2}}\sin 4\unicode[STIX]{x1D719}\,(f_{2}^{\prime \prime \prime }+f_{2}^{\prime \prime })-\cos 4\unicode[STIX]{x1D719}\,f_{1}^{\prime \prime })=0,\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & & \displaystyle (1+{\textstyle \frac{1}{4}}\unicode[STIX]{x1D707}_{2}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3})(f_{2}^{\prime \prime \prime \prime }-2f_{2}^{\prime \prime }+f_{2})+\unicode[STIX]{x1D707}_{1}(\cos 2\unicode[STIX]{x1D719}\,(g_{2}+g_{2}^{\prime \prime })+2\sin 2\unicode[STIX]{x1D719}\,g_{1}^{\prime })\nonumber\\ \displaystyle & & \displaystyle \quad -\,\unicode[STIX]{x1D707}_{2}({\textstyle \frac{1}{2}}\sin 4\unicode[STIX]{x1D719}\,(f_{1}^{\prime \prime \prime }+f_{1}^{\prime \prime })+\cos 4\unicode[STIX]{x1D719}\,f_{2}^{\prime \prime })=0,\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & & \displaystyle (1+{\textstyle \frac{1}{4}}\unicode[STIX]{x1D707}_{2}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3})(f_{2}^{\prime \prime \prime \prime }-2f_{2}^{\prime \prime }+f_{2})+\unicode[STIX]{x1D707}_{1}(\cos 2\unicode[STIX]{x1D719}\,(g_{2}+g_{2}^{\prime \prime })+2\sin 2\unicode[STIX]{x1D719}\,g_{1}^{\prime })\nonumber\\ \displaystyle & & \displaystyle \quad -\,\unicode[STIX]{x1D707}_{2}({\textstyle \frac{1}{2}}\sin 4\unicode[STIX]{x1D719}\,(f_{1}^{\prime \prime \prime }+f_{1}^{\prime \prime })+\cos 4\unicode[STIX]{x1D719}\,f_{2}^{\prime \prime })=0,\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle g_{1}-\sin 2\unicode[STIX]{x1D719}\,f_{1}^{\prime }+\sin ^{2}\!\unicode[STIX]{x1D719}\,f_{2}^{\prime \prime }-\cos ^{2}\!\unicode[STIX]{x1D719}\,f_{2}=0, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle g_{1}-\sin 2\unicode[STIX]{x1D719}\,f_{1}^{\prime }+\sin ^{2}\!\unicode[STIX]{x1D719}\,f_{2}^{\prime \prime }-\cos ^{2}\!\unicode[STIX]{x1D719}\,f_{2}=0, & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle g_{2}-\sin 2\unicode[STIX]{x1D719}\,f_{2}^{\prime }-\sin ^{2}\!\unicode[STIX]{x1D719}\,f_{1}^{\prime \prime }+\cos ^{2}\!\unicode[STIX]{x1D719}\,f_{1}=0, & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle g_{2}-\sin 2\unicode[STIX]{x1D719}\,f_{2}^{\prime }-\sin ^{2}\!\unicode[STIX]{x1D719}\,f_{1}^{\prime \prime }+\cos ^{2}\!\unicode[STIX]{x1D719}\,f_{1}=0, & \displaystyle\end{eqnarray}$$
 $y$
.
$y$
.
Substituting (3.17) and (3.18) into (3.15) and (3.16), the system reduces to two ordinary differential equations for
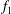 $f_{1}$
and
$f_{1}$
and
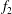 $f_{2}$
. Assuming a basis of solutions of the form
$f_{2}$
. Assuming a basis of solutions of the form
 $$\begin{eqnarray}\left(\begin{array}{@{}c@{}}f_{1}\\ f_{2}\end{array}\right)=\left(\begin{array}{@{}c@{}}f_{1}^{0}\\ f_{2}^{0}\end{array}\right)\text{e}^{\unicode[STIX]{x1D706}y}\end{eqnarray}$$
$$\begin{eqnarray}\left(\begin{array}{@{}c@{}}f_{1}\\ f_{2}\end{array}\right)=\left(\begin{array}{@{}c@{}}f_{1}^{0}\\ f_{2}^{0}\end{array}\right)\text{e}^{\unicode[STIX]{x1D706}y}\end{eqnarray}$$
reduces the problem to the linear system
 $$\begin{eqnarray}\unicode[STIX]{x1D647}\left(\begin{array}{@{}c@{}}f_{1}^{0}\\ f_{2}^{0}\end{array}\right)=\left(\begin{array}{@{}c@{}}0\\ 0\end{array}\right),\quad \text{where }\unicode[STIX]{x1D647}=\left(\begin{array}{@{}cc@{}}\unicode[STIX]{x1D613}_{11} & \unicode[STIX]{x1D613}_{12}\\ \unicode[STIX]{x1D613}_{21} & \unicode[STIX]{x1D613}_{22}\end{array}\right).\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D647}\left(\begin{array}{@{}c@{}}f_{1}^{0}\\ f_{2}^{0}\end{array}\right)=\left(\begin{array}{@{}c@{}}0\\ 0\end{array}\right),\quad \text{where }\unicode[STIX]{x1D647}=\left(\begin{array}{@{}cc@{}}\unicode[STIX]{x1D613}_{11} & \unicode[STIX]{x1D613}_{12}\\ \unicode[STIX]{x1D613}_{21} & \unicode[STIX]{x1D613}_{22}\end{array}\right).\end{eqnarray}$$
The components of
 $\unicode[STIX]{x1D647}$
are given in appendix B. It should be noted that
$\unicode[STIX]{x1D647}$
are given in appendix B. It should be noted that
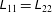 $\unicode[STIX]{x1D613}_{11}=\unicode[STIX]{x1D613}_{22}$
and further that
$\unicode[STIX]{x1D613}_{11}=\unicode[STIX]{x1D613}_{22}$
and further that
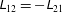 $\unicode[STIX]{x1D613}_{12}=-\unicode[STIX]{x1D613}_{21}$
. For a non-trivial solution, the determinant of the matrix
$\unicode[STIX]{x1D613}_{12}=-\unicode[STIX]{x1D613}_{21}$
. For a non-trivial solution, the determinant of the matrix
 $\unicode[STIX]{x1D647}$
must be zero, yielding the equation
$\unicode[STIX]{x1D647}$
must be zero, yielding the equation
 $$\begin{eqnarray}\unicode[STIX]{x1D613}_{11}^{2}+\unicode[STIX]{x1D613}_{12}^{2}=0;\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D613}_{11}^{2}+\unicode[STIX]{x1D613}_{12}^{2}=0;\end{eqnarray}$$
hence,
 $$\begin{eqnarray}\unicode[STIX]{x1D613}_{11}=\pm \text{i}\unicode[STIX]{x1D613}_{12},\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D613}_{11}=\pm \text{i}\unicode[STIX]{x1D613}_{12},\end{eqnarray}$$
and so
 $$\begin{eqnarray}\displaystyle f_{2}^{0} & = & \displaystyle -\frac{\unicode[STIX]{x1D613}_{11}}{\unicode[STIX]{x1D613}_{12}}f_{1}^{0},\nonumber\\ \displaystyle & = & \displaystyle \mp \text{i}f_{1}^{0}.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle f_{2}^{0} & = & \displaystyle -\frac{\unicode[STIX]{x1D613}_{11}}{\unicode[STIX]{x1D613}_{12}}f_{1}^{0},\nonumber\\ \displaystyle & = & \displaystyle \mp \text{i}f_{1}^{0}.\end{eqnarray}$$
Equation (3.21) has eight complex roots,
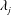 $\unicode[STIX]{x1D706}_{j}$
, four with positive real part and four with negative real part. Since the velocity must remain bounded as
$\unicode[STIX]{x1D706}_{j}$
, four with positive real part and four with negative real part. Since the velocity must remain bounded as
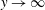 $y\rightarrow \infty$
, we disregard the positive roots. The other four form two complex conjugate pairs,
$y\rightarrow \infty$
, we disregard the positive roots. The other four form two complex conjugate pairs,
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D706}_{1}=\unicode[STIX]{x1D6FC}_{1}+\text{i}\unicode[STIX]{x1D6FD}_{1},\quad \unicode[STIX]{x1D706}_{3}=\unicode[STIX]{x1D6FC}_{1}-\text{i}\unicode[STIX]{x1D6FD}_{1}, & & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D706}_{1}=\unicode[STIX]{x1D6FC}_{1}+\text{i}\unicode[STIX]{x1D6FD}_{1},\quad \unicode[STIX]{x1D706}_{3}=\unicode[STIX]{x1D6FC}_{1}-\text{i}\unicode[STIX]{x1D6FD}_{1}, & & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D706}_{2}=\unicode[STIX]{x1D6FC}_{2}+\text{i}\unicode[STIX]{x1D6FD}_{2},\quad \unicode[STIX]{x1D706}_{4}=\unicode[STIX]{x1D6FC}_{2}-\text{i}\unicode[STIX]{x1D6FD}_{2}. & & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D706}_{2}=\unicode[STIX]{x1D6FC}_{2}+\text{i}\unicode[STIX]{x1D6FD}_{2},\quad \unicode[STIX]{x1D706}_{4}=\unicode[STIX]{x1D6FC}_{2}-\text{i}\unicode[STIX]{x1D6FD}_{2}. & & \displaystyle\end{eqnarray}$$
It should be noted that the
 $\unicode[STIX]{x1D706}_{j}$
are known analytically; however, they are not given here due to space constraints.
$\unicode[STIX]{x1D706}_{j}$
are known analytically; however, they are not given here due to space constraints.
The solution form for
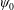 $\unicode[STIX]{x1D713}_{0}$
is thus
$\unicode[STIX]{x1D713}_{0}$
is thus
 $$\begin{eqnarray}\unicode[STIX]{x1D713}_{0}=\mathop{\sum }_{j=1}^{4}\hat{A_{j}}(\cos (x-t)+\unicode[STIX]{x1D709}_{j}\sin (x-t))\text{e}^{\unicode[STIX]{x1D706}_{j}y},\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D713}_{0}=\mathop{\sum }_{j=1}^{4}\hat{A_{j}}(\cos (x-t)+\unicode[STIX]{x1D709}_{j}\sin (x-t))\text{e}^{\unicode[STIX]{x1D706}_{j}y},\end{eqnarray}$$
where
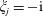 $\unicode[STIX]{x1D709}_{j}=-\text{i}$
for
$\unicode[STIX]{x1D709}_{j}=-\text{i}$
for
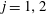 $j=1,2$
and
$j=1,2$
and
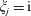 $\unicode[STIX]{x1D709}_{j}=\text{i}$
for
$\unicode[STIX]{x1D709}_{j}=\text{i}$
for
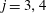 $j=3,4$
. Assuming that the constants take the general form
$j=3,4$
. Assuming that the constants take the general form
 $\hat{A}_{j}=A_{j}+\text{i}B_{j}$
for
$\hat{A}_{j}=A_{j}+\text{i}B_{j}$
for
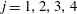 $j=1,2,3,4$
, the boundary conditions (3.12) give
$j=1,2,3,4$
, the boundary conditions (3.12) give
 $$\begin{eqnarray}\displaystyle \left.\begin{array}{@{}c@{}}A_{1}=\displaystyle \frac{\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FD}_{2}-\unicode[STIX]{x1D6FC}_{2}\unicode[STIX]{x1D6FD}_{1}}{2((\unicode[STIX]{x1D6FC}_{1}-\unicode[STIX]{x1D6FC}_{2})^{2}+(\unicode[STIX]{x1D6FD}_{1}-\unicode[STIX]{x1D6FD}_{2})^{2})},\quad A_{2}=-A_{1},\quad A_{3}=A_{1},\quad A_{4}=-A_{1},\\ B_{1}=\displaystyle \frac{\unicode[STIX]{x1D6FC}_{2}^{2}-\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}+\unicode[STIX]{x1D6FD}_{2}^{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2}}{2((\unicode[STIX]{x1D6FC}_{1}-\unicode[STIX]{x1D6FC}_{2})^{2}+(\unicode[STIX]{x1D6FD}_{1}-\unicode[STIX]{x1D6FD}_{2})^{2})},\quad B_{3}=-B_{1},\\ B_{2}=\displaystyle \frac{\unicode[STIX]{x1D6FC}_{1}^{2}-\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}+\unicode[STIX]{x1D6FD}_{1}^{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2}}{2((\unicode[STIX]{x1D6FC}_{1}-\unicode[STIX]{x1D6FC}_{2})^{2}+(\unicode[STIX]{x1D6FD}_{1}-\unicode[STIX]{x1D6FD}_{2})^{2})},\quad B_{4}=-B_{2}.\end{array}\right\} & & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \left.\begin{array}{@{}c@{}}A_{1}=\displaystyle \frac{\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FD}_{2}-\unicode[STIX]{x1D6FC}_{2}\unicode[STIX]{x1D6FD}_{1}}{2((\unicode[STIX]{x1D6FC}_{1}-\unicode[STIX]{x1D6FC}_{2})^{2}+(\unicode[STIX]{x1D6FD}_{1}-\unicode[STIX]{x1D6FD}_{2})^{2})},\quad A_{2}=-A_{1},\quad A_{3}=A_{1},\quad A_{4}=-A_{1},\\ B_{1}=\displaystyle \frac{\unicode[STIX]{x1D6FC}_{2}^{2}-\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}+\unicode[STIX]{x1D6FD}_{2}^{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2}}{2((\unicode[STIX]{x1D6FC}_{1}-\unicode[STIX]{x1D6FC}_{2})^{2}+(\unicode[STIX]{x1D6FD}_{1}-\unicode[STIX]{x1D6FD}_{2})^{2})},\quad B_{3}=-B_{1},\\ B_{2}=\displaystyle \frac{\unicode[STIX]{x1D6FC}_{1}^{2}-\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}+\unicode[STIX]{x1D6FD}_{1}^{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2}}{2((\unicode[STIX]{x1D6FC}_{1}-\unicode[STIX]{x1D6FC}_{2})^{2}+(\unicode[STIX]{x1D6FD}_{1}-\unicode[STIX]{x1D6FD}_{2})^{2})},\quad B_{4}=-B_{2}.\end{array}\right\} & & \displaystyle\end{eqnarray}$$
The fibre angle perturbation is then of the form
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D703}_{0} & = & \displaystyle \mathop{\sum }_{j=1}^{4}\hat{A}_{j}\left[(\unicode[STIX]{x1D706}_{j}\sin 2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D709}_{j}(-\unicode[STIX]{x1D706}_{j}^{2}\sin ^{2}\unicode[STIX]{x1D719}+\cos ^{2}\unicode[STIX]{x1D719}))\cos (x-t)\right.\nonumber\\ \displaystyle & & \displaystyle \left.+\,(\unicode[STIX]{x1D709}_{j}\unicode[STIX]{x1D706}_{j}\sin 2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D706}_{j}^{2}\sin ^{2}\unicode[STIX]{x1D719}-\cos ^{2}\unicode[STIX]{x1D719})\sin (x-t)\right]\text{e}^{\unicode[STIX]{x1D706}_{j}y}.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D703}_{0} & = & \displaystyle \mathop{\sum }_{j=1}^{4}\hat{A}_{j}\left[(\unicode[STIX]{x1D706}_{j}\sin 2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D709}_{j}(-\unicode[STIX]{x1D706}_{j}^{2}\sin ^{2}\unicode[STIX]{x1D719}+\cos ^{2}\unicode[STIX]{x1D719}))\cos (x-t)\right.\nonumber\\ \displaystyle & & \displaystyle \left.+\,(\unicode[STIX]{x1D709}_{j}\unicode[STIX]{x1D706}_{j}\sin 2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D706}_{j}^{2}\sin ^{2}\unicode[STIX]{x1D719}-\cos ^{2}\unicode[STIX]{x1D719})\sin (x-t)\right]\text{e}^{\unicode[STIX]{x1D706}_{j}y}.\end{eqnarray}$$
The change in the small perturbation to the orientation,
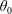 $\unicode[STIX]{x1D703}_{0}$
, is dependent on the initial orientation angle of the fibres along with their position.
$\unicode[STIX]{x1D703}_{0}$
, is dependent on the initial orientation angle of the fibres along with their position.
3.4 Order-
 $\unicode[STIX]{x1D700}^{2}$
solution and mean swimming velocity
$\unicode[STIX]{x1D700}^{2}$
solution and mean swimming velocity

The mean swimming velocity is determined by the horizontal component of the flow as
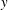 $y$
tends to infinity. The leading-order stream function, equation (3.26), tends to zero, and hence the non-zero mean swimming velocity is determined at order
$y$
tends to infinity. The leading-order stream function, equation (3.26), tends to zero, and hence the non-zero mean swimming velocity is determined at order
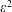 $\unicode[STIX]{x1D700}^{2}$
,
$\unicode[STIX]{x1D700}^{2}$
,
 $$\begin{eqnarray}U\sim \unicode[STIX]{x1D700}^{2}U_{1}=\lim _{y\rightarrow \infty }\unicode[STIX]{x1D700}^{2}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}y^{}}}.\end{eqnarray}$$
$$\begin{eqnarray}U\sim \unicode[STIX]{x1D700}^{2}U_{1}=\lim _{y\rightarrow \infty }\unicode[STIX]{x1D700}^{2}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}y^{}}}.\end{eqnarray}$$
We neglect the oscillatory terms to determine the leading-order term in the expansion of mean swimming velocity, which we denote as
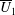 $\overline{U}_{1}$
. The bar notation represents an average over one time period.
$\overline{U}_{1}$
. The bar notation represents an average over one time period.
At order
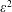 $\unicode[STIX]{x1D700}^{2}$
, the boundary conditions (3.6) and (3.7) become
$\unicode[STIX]{x1D700}^{2}$
, the boundary conditions (3.6) and (3.7) become
 $$\begin{eqnarray}\left.\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}y}\right|_{y=0}+\sin (x-t)\left.\frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{2}}\right|_{y=0}=0,\quad \left.\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x}\right|_{y=0}+\sin (x-t)\left.\frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y\unicode[STIX]{x2202}x}\right|_{y=0}=0,\end{eqnarray}$$
$$\begin{eqnarray}\left.\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}y}\right|_{y=0}+\sin (x-t)\left.\frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{2}}\right|_{y=0}=0,\quad \left.\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x}\right|_{y=0}+\sin (x-t)\left.\frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y\unicode[STIX]{x2202}x}\right|_{y=0}=0,\end{eqnarray}$$
and hence
 $$\begin{eqnarray}\displaystyle & \displaystyle \left.{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}y^{}}}\right|_{y=0}=\frac{1}{2}\left((\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2})(1-\cos 2(x-t))-(\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FD}_{2}-\unicode[STIX]{x1D6FC}_{2}\unicode[STIX]{x1D6FD}_{1})\sin 2(x-t)\right),\qquad & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \left.{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}y^{}}}\right|_{y=0}=\frac{1}{2}\left((\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2})(1-\cos 2(x-t))-(\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FD}_{2}-\unicode[STIX]{x1D6FC}_{2}\unicode[STIX]{x1D6FD}_{1})\sin 2(x-t)\right),\qquad & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle \left.{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x^{}}}\right|_{y=0}=0. & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \left.{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x^{}}}\right|_{y=0}=0. & \displaystyle\end{eqnarray}$$
Because of the form of the boundary conditions, the ansatz is
 $$\begin{eqnarray}\unicode[STIX]{x1D713}_{1}=\hat{f}_{1}(y)+\hat{f}_{2}(y)\cos 2(x-t)+\hat{f}_{3}(y)\sin 2(x-t),\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D713}_{1}=\hat{f}_{1}(y)+\hat{f}_{2}(y)\cos 2(x-t)+\hat{f}_{3}(y)\sin 2(x-t),\end{eqnarray}$$
for some functions
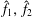 $\hat{f}_{1},\,\hat{f}_{2}$
and
$\hat{f}_{1},\,\hat{f}_{2}$
and
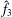 $\hat{f}_{3}$
. Substituting the solution form (3.33) into the order-
$\hat{f}_{3}$
. Substituting the solution form (3.33) into the order-
 $\unicode[STIX]{x1D700}^{2}$
expansion of (3.3), shown in full in appendix B, and equating coefficients of non-oscillating terms, we have
$\unicode[STIX]{x1D700}^{2}$
expansion of (3.3), shown in full in appendix B, and equating coefficients of non-oscillating terms, we have
 $$\begin{eqnarray}\hat{f}_{1}^{\prime \prime \prime \prime }=0,\end{eqnarray}$$
$$\begin{eqnarray}\hat{f}_{1}^{\prime \prime \prime \prime }=0,\end{eqnarray}$$
and hence
 $$\begin{eqnarray}\hat{f}_{1}(y)=Ay^{3}+By^{2}+Cy+D.\end{eqnarray}$$
$$\begin{eqnarray}\hat{f}_{1}(y)=Ay^{3}+By^{2}+Cy+D.\end{eqnarray}$$
To ensure that the velocity remains bounded as
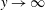 $y\rightarrow \infty$
, we set
$y\rightarrow \infty$
, we set
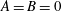 $A=B=0$
. From the boundary conditions (3.31) and (3.32), we find that
$A=B=0$
. From the boundary conditions (3.31) and (3.32), we find that
 $C=(\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2})/2$
and
$C=(\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2})/2$
and
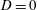 $D=0$
, and hence
$D=0$
, and hence
 $$\begin{eqnarray}\hat{f}_{1}(y)=\frac{y}{2}(\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2}).\end{eqnarray}$$
$$\begin{eqnarray}\hat{f}_{1}(y)=\frac{y}{2}(\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2}).\end{eqnarray}$$
Differentiating
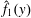 $\hat{f}_{1}(y)$
with respect to
$\hat{f}_{1}(y)$
with respect to
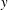 $y$
, the leading-order term in the expansion of mean swimming velocity is calculated as
$y$
, the leading-order term in the expansion of mean swimming velocity is calculated as
 $$\begin{eqnarray}\overline{U}_{1}={\textstyle \frac{1}{2}}(\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2}).\end{eqnarray}$$
$$\begin{eqnarray}\overline{U}_{1}={\textstyle \frac{1}{2}}(\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FD}_{1}\unicode[STIX]{x1D6FD}_{2}).\end{eqnarray}$$
3.5 Mean rate of working
To determine how Stokesian swimming is affected by transverse isotropy, the mean rate of working at order
 $\unicode[STIX]{x1D700}$
is investigated, i.e. the rate of working per unit area of the sheet against viscous stress,
$\unicode[STIX]{x1D700}$
is investigated, i.e. the rate of working per unit area of the sheet against viscous stress,
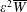 $\unicode[STIX]{x1D700}^{2}\overline{W}$
(Taylor Reference Taylor1951). The mean value of this quantity is given by
$\unicode[STIX]{x1D700}^{2}\overline{W}$
(Taylor Reference Taylor1951). The mean value of this quantity is given by
 $$\begin{eqnarray}\unicode[STIX]{x1D700}^{2}\overline{W}=-\overline{\left.\frac{\unicode[STIX]{x2202}y_{s}}{\unicode[STIX]{x2202}t}\unicode[STIX]{x1D70E}_{22}\right|_{y=0}},\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D700}^{2}\overline{W}=-\overline{\left.\frac{\unicode[STIX]{x2202}y_{s}}{\unicode[STIX]{x2202}t}\unicode[STIX]{x1D70E}_{22}\right|_{y=0}},\end{eqnarray}$$
where
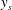 $y_{s}$
is the equation of the sheet surface and
$y_{s}$
is the equation of the sheet surface and
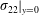 $\unicode[STIX]{x1D70E}_{22}|_{y=0}$
is the normal stress evaluated on the sheet. The no-slip condition is
$\unicode[STIX]{x1D70E}_{22}|_{y=0}$
is the normal stress evaluated on the sheet. The no-slip condition is
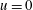 $u=0$
on the sheet, and hence
$u=0$
on the sheet, and hence
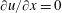 $\unicode[STIX]{x2202}u/\unicode[STIX]{x2202}x=0$
, and, via (2.8),
$\unicode[STIX]{x2202}u/\unicode[STIX]{x2202}x=0$
, and, via (2.8),
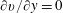 $\unicode[STIX]{x2202}v/\unicode[STIX]{x2202}y=0$
. In terms of the stream function,
$\unicode[STIX]{x2202}v/\unicode[STIX]{x2202}y=0$
. In terms of the stream function,
 $\unicode[STIX]{x1D70E}_{22}$
is
$\unicode[STIX]{x1D70E}_{22}$
is
 $$\begin{eqnarray}\unicode[STIX]{x1D70E}_{22}=-p+\unicode[STIX]{x1D707}_{1}\sin ^{2}\unicode[STIX]{x1D719}+(\unicode[STIX]{x1D707}_{2}\cos \unicode[STIX]{x1D719}\sin ^{3}\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\sin 2\unicode[STIX]{x1D719})\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\right).\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D70E}_{22}=-p+\unicode[STIX]{x1D707}_{1}\sin ^{2}\unicode[STIX]{x1D719}+(\unicode[STIX]{x1D707}_{2}\cos \unicode[STIX]{x1D719}\sin ^{3}\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\sin 2\unicode[STIX]{x1D719})\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}}{\unicode[STIX]{x2202}x^{2}}}\right).\end{eqnarray}$$
Solving (2.9), using the leading-order expression for the stream function (3.26), determines the pressure. Noting that
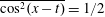 $\overline{\cos ^{2}(x-t)}=1/2$
, we obtain an expression for the leading-order term in the expansion of the mean rate of working,
$\overline{\cos ^{2}(x-t)}=1/2$
, we obtain an expression for the leading-order term in the expansion of the mean rate of working,
 $$\begin{eqnarray}\displaystyle \overline{W} & = & \displaystyle -{\textstyle \frac{1}{16}} [\!(\unicode[STIX]{x1D6FC}_{1}^{2}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FC}_{2}\unicode[STIX]{x1D6FD}_{1}(\unicode[STIX]{x1D6FD}_{1}+2\unicode[STIX]{x1D6FD}_{2})\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D6FC}_{1}(\unicode[STIX]{x1D6FC}_{2}^{2}-\unicode[STIX]{x1D6FD}_{2}(2\unicode[STIX]{x1D6FD}_{1}+\unicode[STIX]{x1D6FD}_{2}))\!)(8+\unicode[STIX]{x1D707}_{2}(1+\cos 4\unicode[STIX]{x1D719})+8\unicode[STIX]{x1D707}_{3})\nonumber\\ \displaystyle & & \displaystyle +\,4(\unicode[STIX]{x1D6FC}_{2}\unicode[STIX]{x1D6FD}_{1}+\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FD}_{2})\unicode[STIX]{x1D707}_{2}\sin 4\unicode[STIX]{x1D719}\!].\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \overline{W} & = & \displaystyle -{\textstyle \frac{1}{16}} [\!(\unicode[STIX]{x1D6FC}_{1}^{2}\unicode[STIX]{x1D6FC}_{2}-\unicode[STIX]{x1D6FC}_{2}\unicode[STIX]{x1D6FD}_{1}(\unicode[STIX]{x1D6FD}_{1}+2\unicode[STIX]{x1D6FD}_{2})\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D6FC}_{1}(\unicode[STIX]{x1D6FC}_{2}^{2}-\unicode[STIX]{x1D6FD}_{2}(2\unicode[STIX]{x1D6FD}_{1}+\unicode[STIX]{x1D6FD}_{2}))\!)(8+\unicode[STIX]{x1D707}_{2}(1+\cos 4\unicode[STIX]{x1D719})+8\unicode[STIX]{x1D707}_{3})\nonumber\\ \displaystyle & & \displaystyle +\,4(\unicode[STIX]{x1D6FC}_{2}\unicode[STIX]{x1D6FD}_{1}+\unicode[STIX]{x1D6FC}_{1}\unicode[STIX]{x1D6FD}_{2})\unicode[STIX]{x1D707}_{2}\sin 4\unicode[STIX]{x1D719}\!].\end{eqnarray}$$
4 Results
The leading-order terms in the expansions of the mean swimming velocity (3.37), the mean rate of working (3.40), the fibre perturbation (3.28), the velocities
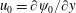 $u_{0}=\unicode[STIX]{x2202}\unicode[STIX]{x1D713}_{0}/\unicode[STIX]{x2202}y$
,
$u_{0}=\unicode[STIX]{x2202}\unicode[STIX]{x1D713}_{0}/\unicode[STIX]{x2202}y$
,
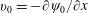 $v_{0}=-\unicode[STIX]{x2202}\unicode[STIX]{x1D713}_{0}/\unicode[STIX]{x2202}x$
and the stream function (3.26) have been found analytically in terms of lengthy expressions for
$v_{0}=-\unicode[STIX]{x2202}\unicode[STIX]{x1D713}_{0}/\unicode[STIX]{x2202}x$
and the stream function (3.26) have been found analytically in terms of lengthy expressions for
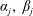 $\unicode[STIX]{x1D6FC}_{j},\,\unicode[STIX]{x1D6FD}_{j}$
. The analytical results for the mean rate of working have been recreated numerically using finite differences and integration by the midpoint method. Each separate component has been verified along with the full solution. The solutions agreed to within a small degree of numerical error. For brevity, we will refer to the time averages of the leading-order terms in the expressions for the swimming velocity and the rate of working as the mean swimming velocity and the mean rate of working respectively, and we will plot terms without the leading
$\unicode[STIX]{x1D6FC}_{j},\,\unicode[STIX]{x1D6FD}_{j}$
. The analytical results for the mean rate of working have been recreated numerically using finite differences and integration by the midpoint method. Each separate component has been verified along with the full solution. The solutions agreed to within a small degree of numerical error. For brevity, we will refer to the time averages of the leading-order terms in the expressions for the swimming velocity and the rate of working as the mean swimming velocity and the mean rate of working respectively, and we will plot terms without the leading
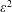 $\unicode[STIX]{x1D700}^{2}$
factors as defined by
$\unicode[STIX]{x1D700}^{2}$
factors as defined by
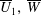 $\overline{U}_{1},\,\overline{W}$
in (3.29) and (3.38).
$\overline{U}_{1},\,\overline{W}$
in (3.29) and (3.38).
We now discuss the results in more detail. Four different flow regimes are considered (figure 3): (i) a passive transversely isotropic fluid, occurring when
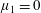 $\unicode[STIX]{x1D707}_{1}=0$
; (ii) an active fluid where
$\unicode[STIX]{x1D707}_{1}=0$
; (ii) an active fluid where
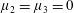 $\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
and
$\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
and
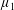 $\unicode[STIX]{x1D707}_{1}$
is non-zero; (iii) a nearly isotropic regime, where all parameters take values up to
$\unicode[STIX]{x1D707}_{1}$
is non-zero; (iii) a nearly isotropic regime, where all parameters take values up to
 $5$
; (iv) the regime where at least one of
$5$
; (iv) the regime where at least one of
 $\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
 $\unicode[STIX]{x1D707}_{3}$
is much larger than 1. A range of initial orientation angles,
$\unicode[STIX]{x1D707}_{3}$
is much larger than 1. A range of initial orientation angles,
 $\unicode[STIX]{x1D719}$
, between
$\unicode[STIX]{x1D719}$
, between
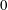 $0$
and
$0$
and
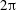 $2\unicode[STIX]{x03C0}$
is considered for all regimes and the active parameter,
$2\unicode[STIX]{x03C0}$
is considered for all regimes and the active parameter,
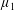 $\unicode[STIX]{x1D707}_{1}$
, is allowed to take both positive and negative values to account for ‘puller’ and ‘pusher’ active behaviour respectively. It should be noted that since the fibres have no directionality, the regime
$\unicode[STIX]{x1D707}_{1}$
, is allowed to take both positive and negative values to account for ‘puller’ and ‘pusher’ active behaviour respectively. It should be noted that since the fibres have no directionality, the regime
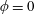 $\unicode[STIX]{x1D719}=0$
–
$\unicode[STIX]{x1D719}=0$
–
 $\unicode[STIX]{x03C0}$
is identical to
$\unicode[STIX]{x03C0}$
is identical to
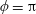 $\unicode[STIX]{x1D719}=\unicode[STIX]{x03C0}$
–
$\unicode[STIX]{x1D719}=\unicode[STIX]{x03C0}$
–
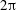 $2\unicode[STIX]{x03C0}$
.
$2\unicode[STIX]{x03C0}$
.
4.1 Regime (i): effect of passive fibres on mean swimming velocity and rate of working
Passive fibres exert no shear-independent force and have no self-propulsive properties; hence, the active parameter
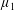 $\unicode[STIX]{x1D707}_{1}$
is set to zero. In this regime, the mean swimming velocity takes on the Newtonian value,
$\unicode[STIX]{x1D707}_{1}$
is set to zero. In this regime, the mean swimming velocity takes on the Newtonian value,
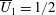 $\overline{U}_{1}=1/2$
, and the mean rate of working is independent of the initial orientation angle
$\overline{U}_{1}=1/2$
, and the mean rate of working is independent of the initial orientation angle
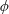 $\unicode[STIX]{x1D719}$
. In figure 5(a), the mean rate of working is always greater than or equal to the Newtonian case,
$\unicode[STIX]{x1D719}$
. In figure 5(a), the mean rate of working is always greater than or equal to the Newtonian case,
 $\overline{W}=1$
. The increase in the mean rate of working is linear throughout, with
$\overline{W}=1$
. The increase in the mean rate of working is linear throughout, with
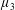 $\unicode[STIX]{x1D707}_{3}$
having a larger impact than
$\unicode[STIX]{x1D707}_{3}$
having a larger impact than
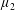 $\unicode[STIX]{x1D707}_{2}$
. Figure 5(b) depicts the relationship between the mean rate of working and the parallel viscosity
$\unicode[STIX]{x1D707}_{2}$
. Figure 5(b) depicts the relationship between the mean rate of working and the parallel viscosity
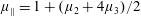 $\unicode[STIX]{x1D707}_{\Vert }=1+(\unicode[STIX]{x1D707}_{2}+4\unicode[STIX]{x1D707}_{3})/2$
, where each line represents a different value of
$\unicode[STIX]{x1D707}_{\Vert }=1+(\unicode[STIX]{x1D707}_{2}+4\unicode[STIX]{x1D707}_{3})/2$
, where each line represents a different value of
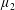 $\unicode[STIX]{x1D707}_{2}$
. The increase in the mean rate of working with
$\unicode[STIX]{x1D707}_{2}$
. The increase in the mean rate of working with
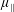 $\unicode[STIX]{x1D707}_{\Vert }$
is linear, apart from the case where
$\unicode[STIX]{x1D707}_{\Vert }$
is linear, apart from the case where
 $\unicode[STIX]{x1D707}_{3}$
is small and
$\unicode[STIX]{x1D707}_{3}$
is small and
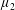 $\unicode[STIX]{x1D707}_{2}$
is large, with a large mean rate of working as
$\unicode[STIX]{x1D707}_{2}$
is large, with a large mean rate of working as
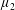 $\unicode[STIX]{x1D707}_{2}$
increases.
$\unicode[STIX]{x1D707}_{2}$
increases.

Figure 5. Mean rate of working by the swimmer in a transversely isotropic fluid where
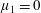 $\unicode[STIX]{x1D707}_{1}=0$
. (a) The mean rate of working for varying
$\unicode[STIX]{x1D707}_{1}=0$
. (a) The mean rate of working for varying
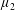 $\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{2}$
and
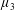 $\unicode[STIX]{x1D707}_{3}$
. This result is identical for all initial angles
$\unicode[STIX]{x1D707}_{3}$
. This result is identical for all initial angles
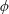 $\unicode[STIX]{x1D719}$
. (b) The change in the mean rate of working for increasing
$\unicode[STIX]{x1D719}$
. (b) The change in the mean rate of working for increasing
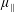 $\unicode[STIX]{x1D707}_{\Vert }$
for a range of
$\unicode[STIX]{x1D707}_{\Vert }$
for a range of
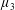 $\unicode[STIX]{x1D707}_{3}$
and set
$\unicode[STIX]{x1D707}_{3}$
and set
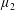 $\unicode[STIX]{x1D707}_{2}$
values:
$\unicode[STIX]{x1D707}_{2}$
values:
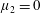 $\unicode[STIX]{x1D707}_{2}=0$
(solid line),
$\unicode[STIX]{x1D707}_{2}=0$
(solid line),
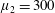 $\unicode[STIX]{x1D707}_{2}=300$
(dashed line),
$\unicode[STIX]{x1D707}_{2}=300$
(dashed line),
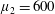 $\unicode[STIX]{x1D707}_{2}=600$
(dash-dotted line) and
$\unicode[STIX]{x1D707}_{2}=600$
(dash-dotted line) and
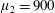 $\unicode[STIX]{x1D707}_{2}=900$
(dotted line).
$\unicode[STIX]{x1D707}_{2}=900$
(dotted line).
4.2 Regime (ii): active-only effects on mean swimming velocity and rate of working
The active-only regime considers
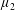 $\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{2}$
and
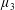 $\unicode[STIX]{x1D707}_{3}$
to be zero with
$\unicode[STIX]{x1D707}_{3}$
to be zero with
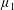 $\unicode[STIX]{x1D707}_{1}$
non-zero. The mean swimming velocity is considered in figure 6 and the mean rate of working in figure 7. For
$\unicode[STIX]{x1D707}_{1}$
non-zero. The mean swimming velocity is considered in figure 6 and the mean rate of working in figure 7. For
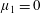 $\unicode[STIX]{x1D707}_{1}=0$
, we regain the Newtonian result, and hence both the mean swimming velocity and the mean rate of working are independent of the fibre angle. For non-zero active parameter
$\unicode[STIX]{x1D707}_{1}=0$
, we regain the Newtonian result, and hence both the mean swimming velocity and the mean rate of working are independent of the fibre angle. For non-zero active parameter
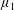 $\unicode[STIX]{x1D707}_{1}$
, the mean swimming velocity and mean rate of working vary considerably with the fibre angle. In particular, negative mean swimming velocity – i.e. reversal of swimming direction – and negative mean rate of working are observed in certain regimes for large
$\unicode[STIX]{x1D707}_{1}$
, the mean swimming velocity and mean rate of working vary considerably with the fibre angle. In particular, negative mean swimming velocity – i.e. reversal of swimming direction – and negative mean rate of working are observed in certain regimes for large
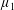 $\unicode[STIX]{x1D707}_{1}$
, with a sudden and dramatic switch in sign close to
$\unicode[STIX]{x1D707}_{1}$
, with a sudden and dramatic switch in sign close to
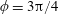 $\unicode[STIX]{x1D719}=3\unicode[STIX]{x03C0}/4$
(figures 6
c,d and 7
c,d). It should be noted that this can be resolved through refinement of the plotting grid and is not a discontinuity. A change from ‘pusher’- to ‘puller’-type active behaviour (equivalent to a change in sign of
$\unicode[STIX]{x1D719}=3\unicode[STIX]{x03C0}/4$
(figures 6
c,d and 7
c,d). It should be noted that this can be resolved through refinement of the plotting grid and is not a discontinuity. A change from ‘pusher’- to ‘puller’-type active behaviour (equivalent to a change in sign of
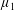 $\unicode[STIX]{x1D707}_{1}$
) is equivalent to a reflection in the line
$\unicode[STIX]{x1D707}_{1}$
) is equivalent to a reflection in the line
 $\unicode[STIX]{x1D719}=\unicode[STIX]{x03C0}/2$
(
$\unicode[STIX]{x1D719}=\unicode[STIX]{x03C0}/2$
(
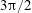 $3\unicode[STIX]{x03C0}/2$
).
$3\unicode[STIX]{x03C0}/2$
).

Figure 6. Mean swimming velocity versus the initial orientation
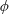 $\unicode[STIX]{x1D719}$
, where
$\unicode[STIX]{x1D719}$
, where
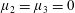 $\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
. Panels (a) and (b) depict small positive and negative
$\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
. Panels (a) and (b) depict small positive and negative
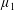 $\unicode[STIX]{x1D707}_{1}$
values:
$\unicode[STIX]{x1D707}_{1}$
values:
 $0$
(solid line),
$0$
(solid line),
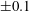 $\pm 0.1$
(dashed line),
$\pm 0.1$
(dashed line),
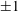 $\pm 1$
(dash-dotted line) and
$\pm 1$
(dash-dotted line) and
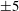 $\pm 5$
(dotted line). Panels (c) and (d) depict larger
$\pm 5$
(dotted line). Panels (c) and (d) depict larger
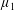 $\unicode[STIX]{x1D707}_{1}$
values:
$\unicode[STIX]{x1D707}_{1}$
values:
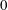 $0$
(solid line),
$0$
(solid line),
 $\pm 300$
(dashed line),
$\pm 300$
(dashed line),
 $\pm 600$
(dash-dotted line),
$\pm 600$
(dash-dotted line),
 $\pm 900$
(dotted line), where the arrows denote increasing
$\pm 900$
(dotted line), where the arrows denote increasing
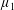 $\unicode[STIX]{x1D707}_{1}$
.
$\unicode[STIX]{x1D707}_{1}$
.

Figure 7. Mean rate of working versus the initial orientation
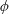 $\unicode[STIX]{x1D719}$
, where
$\unicode[STIX]{x1D719}$
, where
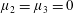 $\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
. Panels (a) and (b) depict small positive and negative
$\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
. Panels (a) and (b) depict small positive and negative
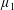 $\unicode[STIX]{x1D707}_{1}$
values:
$\unicode[STIX]{x1D707}_{1}$
values:
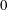 $0$
(solid line),
$0$
(solid line),
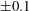 $\pm 0.1$
(dashed line),
$\pm 0.1$
(dashed line),
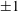 $\pm 1$
(dash-dotted line) and
$\pm 1$
(dash-dotted line) and
 $\pm 5$
(dotted line). Panels (c) and (d) depict larger
$\pm 5$
(dotted line). Panels (c) and (d) depict larger
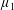 $\unicode[STIX]{x1D707}_{1}$
values:
$\unicode[STIX]{x1D707}_{1}$
values:
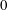 $0$
(solid line),
$0$
(solid line),
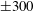 $\pm 300$
(dashed line),
$\pm 300$
(dashed line),
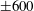 $\pm 600$
(dash-dotted line),
$\pm 600$
(dash-dotted line),
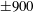 $\pm 900$
(dotted line), where the arrows denote increasing
$\pm 900$
(dotted line), where the arrows denote increasing
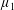 $\unicode[STIX]{x1D707}_{1}$
.
$\unicode[STIX]{x1D707}_{1}$
.
4.3 Regime (iii): nearly isotropic behaviour in leading-order mean swimming velocity and rate of working
A small perturbation away from the isotropic case is considered here;
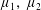 $\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
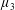 $\unicode[STIX]{x1D707}_{3}$
take values of up to
$\unicode[STIX]{x1D707}_{3}$
take values of up to
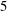 $5$
. When
$5$
. When
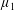 $\unicode[STIX]{x1D707}_{1}$
is much smaller than 1 and positive (figure 8
a,b), a small perturbation away from the Newtonian case is observed. As
$\unicode[STIX]{x1D707}_{1}$
is much smaller than 1 and positive (figure 8
a,b), a small perturbation away from the Newtonian case is observed. As
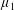 $\unicode[STIX]{x1D707}_{1}$
continues to increase, angular dependence becomes more prevalent. For the mean swimming velocity,
$\unicode[STIX]{x1D707}_{1}$
continues to increase, angular dependence becomes more prevalent. For the mean swimming velocity,
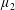 $\unicode[STIX]{x1D707}_{2}$
has minimal impact, while
$\unicode[STIX]{x1D707}_{2}$
has minimal impact, while
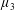 $\unicode[STIX]{x1D707}_{3}$
reduces the range of values that the background flow can take. For the mean rate of working (figure 9),
$\unicode[STIX]{x1D707}_{3}$
reduces the range of values that the background flow can take. For the mean rate of working (figure 9),
 $\unicode[STIX]{x1D707}_{2}$
again has little impact on the results, and the effect of increasing
$\unicode[STIX]{x1D707}_{2}$
again has little impact on the results, and the effect of increasing
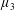 $\unicode[STIX]{x1D707}_{3}$
is to increase the cost of swimming. When
$\unicode[STIX]{x1D707}_{3}$
is to increase the cost of swimming. When
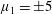 $\unicode[STIX]{x1D707}_{1}=\pm 5$
(figure 9
e,f), the mean rate of working may become negative, and the effect of increasing
$\unicode[STIX]{x1D707}_{1}=\pm 5$
(figure 9
e,f), the mean rate of working may become negative, and the effect of increasing
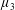 $\unicode[STIX]{x1D707}_{3}$
is to reduce the range of values that the mean rate of working will take.
$\unicode[STIX]{x1D707}_{3}$
is to reduce the range of values that the mean rate of working will take.

Figure 8. Mean swimming velocity versus
 $\unicode[STIX]{x1D719}$
, where the parameters
$\unicode[STIX]{x1D719}$
, where the parameters
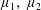 $\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
 $\unicode[STIX]{x1D707}_{3}$
take values of up to
$\unicode[STIX]{x1D707}_{3}$
take values of up to
 $5$
. Panels (a) and (b) show
$5$
. Panels (a) and (b) show
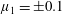 $\unicode[STIX]{x1D707}_{1}=\pm 0.1$
, (c) and (d) show
$\unicode[STIX]{x1D707}_{1}=\pm 0.1$
, (c) and (d) show
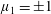 $\unicode[STIX]{x1D707}_{1}=\pm 1$
and (e) and (f) show
$\unicode[STIX]{x1D707}_{1}=\pm 1$
and (e) and (f) show
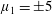 $\unicode[STIX]{x1D707}_{1}=\pm 5$
. Each line depicts a different
$\unicode[STIX]{x1D707}_{1}=\pm 5$
. Each line depicts a different
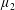 $\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{2}$
and
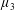 $\unicode[STIX]{x1D707}_{3}$
combination:
$\unicode[STIX]{x1D707}_{3}$
combination:
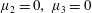 $\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=0$
(solid line),
$\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=0$
(solid line),
 $\unicode[STIX]{x1D707}_{2}=1,\,\unicode[STIX]{x1D707}_{3}=0$
(dashed line),
$\unicode[STIX]{x1D707}_{2}=1,\,\unicode[STIX]{x1D707}_{3}=0$
(dashed line),
 $\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=1$
(dash-dotted line) and
$\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=1$
(dash-dotted line) and
 $\unicode[STIX]{x1D707}_{2}=1,\,\unicode[STIX]{x1D707}_{3}=1$
(dotted line).
$\unicode[STIX]{x1D707}_{2}=1,\,\unicode[STIX]{x1D707}_{3}=1$
(dotted line).
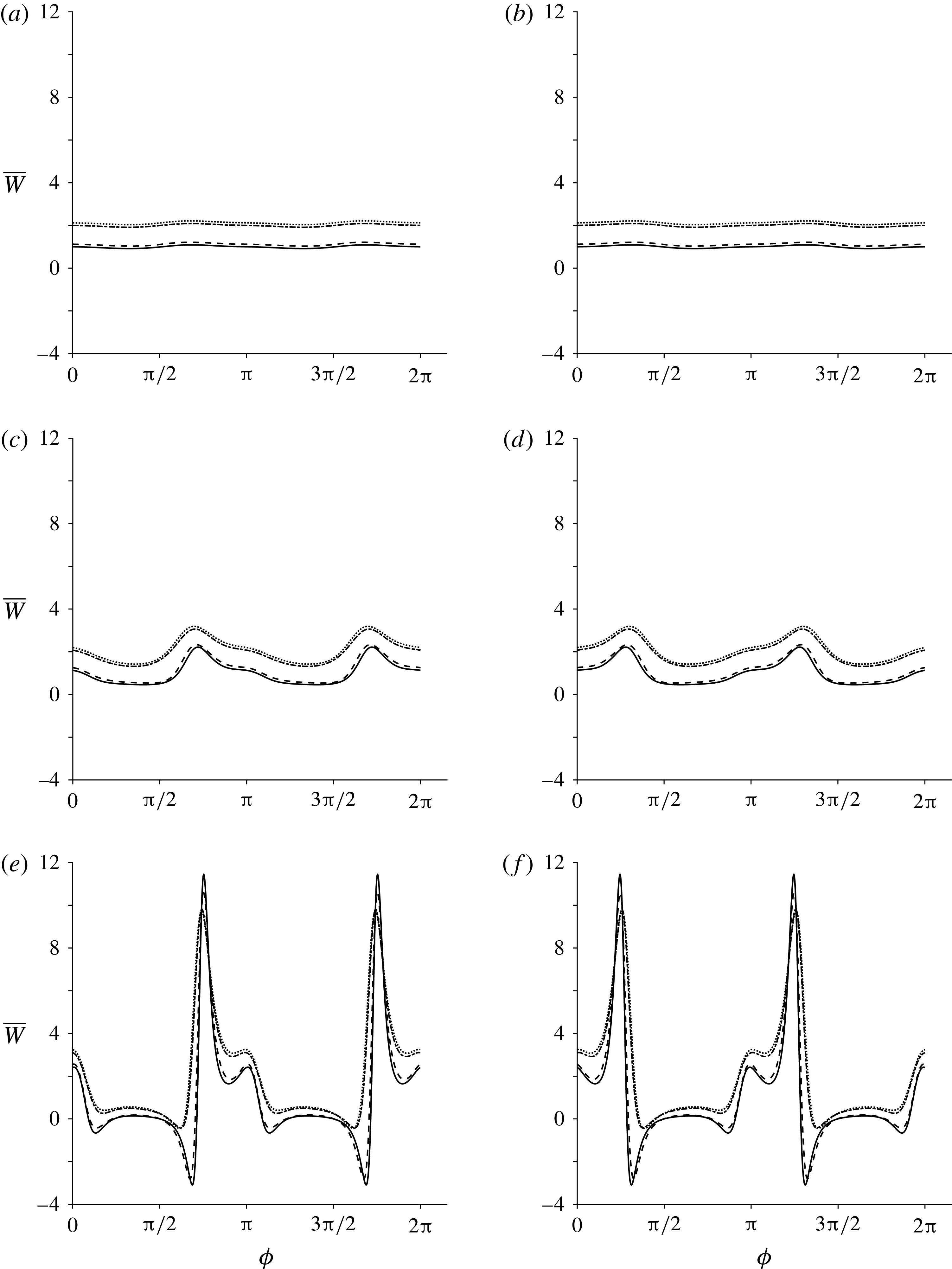
Figure 9. Mean rate of working versus
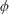 $\unicode[STIX]{x1D719}$
, where the parameters
$\unicode[STIX]{x1D719}$
, where the parameters
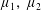 $\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
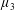 $\unicode[STIX]{x1D707}_{3}$
take values of up to
$\unicode[STIX]{x1D707}_{3}$
take values of up to
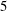 $5$
. Panels (a) and (b) show
$5$
. Panels (a) and (b) show
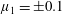 $\unicode[STIX]{x1D707}_{1}=\pm 0.1$
, (c) and (d) show
$\unicode[STIX]{x1D707}_{1}=\pm 0.1$
, (c) and (d) show
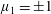 $\unicode[STIX]{x1D707}_{1}=\pm 1$
and (e) and (f) show
$\unicode[STIX]{x1D707}_{1}=\pm 1$
and (e) and (f) show
 $\unicode[STIX]{x1D707}_{1}=\pm 5$
. Each line depicts a different
$\unicode[STIX]{x1D707}_{1}=\pm 5$
. Each line depicts a different
 $\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{2}$
and
 $\unicode[STIX]{x1D707}_{3}$
combination:
$\unicode[STIX]{x1D707}_{3}$
combination:
 $\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=0$
(solid line),
$\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=0$
(solid line),
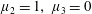 $\unicode[STIX]{x1D707}_{2}=1,\,\unicode[STIX]{x1D707}_{3}=0$
(dashed line),
$\unicode[STIX]{x1D707}_{2}=1,\,\unicode[STIX]{x1D707}_{3}=0$
(dashed line),
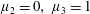 $\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=1$
(dash-dotted line) and
$\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=1$
(dash-dotted line) and
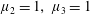 $\unicode[STIX]{x1D707}_{2}=1,\,\unicode[STIX]{x1D707}_{3}=1$
(dotted line).
$\unicode[STIX]{x1D707}_{2}=1,\,\unicode[STIX]{x1D707}_{3}=1$
(dotted line).
4.4 Regime (iv): the effect of large rheological parameters on leading-order mean swimming velocity and rate of working
The final regime is where at least one of
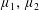 $\unicode[STIX]{x1D707}_{1},\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{1},\unicode[STIX]{x1D707}_{2}$
and
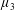 $\unicode[STIX]{x1D707}_{3}$
is much larger than 1. Figures 10 and 11 depict how the mean swimming velocity and mean rate of working change with the initial orientation angle,
$\unicode[STIX]{x1D707}_{3}$
is much larger than 1. Figures 10 and 11 depict how the mean swimming velocity and mean rate of working change with the initial orientation angle,
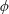 $\unicode[STIX]{x1D719}$
. When either
$\unicode[STIX]{x1D719}$
. When either
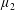 $\unicode[STIX]{x1D707}_{2}$
or
$\unicode[STIX]{x1D707}_{2}$
or
 $\unicode[STIX]{x1D707}_{3}$
is non-zero, the steep peaks that occurred at
$\unicode[STIX]{x1D707}_{3}$
is non-zero, the steep peaks that occurred at
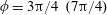 $\unicode[STIX]{x1D719}=3\unicode[STIX]{x03C0}/4~(7\unicode[STIX]{x03C0}/4)$
(figure 6
c) and
$\unicode[STIX]{x1D719}=3\unicode[STIX]{x03C0}/4~(7\unicode[STIX]{x03C0}/4)$
(figure 6
c) and
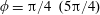 $\unicode[STIX]{x1D719}=\unicode[STIX]{x03C0}/4~(5\unicode[STIX]{x03C0}/4)$
(figure 6
d) within regime (ii) no longer appear. Further, when
$\unicode[STIX]{x1D719}=\unicode[STIX]{x03C0}/4~(5\unicode[STIX]{x03C0}/4)$
(figure 6
d) within regime (ii) no longer appear. Further, when
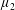 $\unicode[STIX]{x1D707}_{2}$
is non-zero and
$\unicode[STIX]{x1D707}_{2}$
is non-zero and
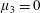 $\unicode[STIX]{x1D707}_{3}=0$
, the mean swimming velocity becomes negative for certain initial orientation angles, i.e. the swimming direction is reversed. When
$\unicode[STIX]{x1D707}_{3}=0$
, the mean swimming velocity becomes negative for certain initial orientation angles, i.e. the swimming direction is reversed. When
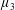 $\unicode[STIX]{x1D707}_{3}$
becomes non-zero, the results collapse down towards the Newtonian case, altered predominantly by the active parameter,
$\unicode[STIX]{x1D707}_{3}$
becomes non-zero, the results collapse down towards the Newtonian case, altered predominantly by the active parameter,
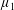 $\unicode[STIX]{x1D707}_{1}$
. Similar results are seen for the mean rate of working; however, for non-zero
$\unicode[STIX]{x1D707}_{1}$
. Similar results are seen for the mean rate of working; however, for non-zero
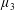 $\unicode[STIX]{x1D707}_{3}$
(figure 11
c,d), the reference value about which variations occur is significantly increased.
$\unicode[STIX]{x1D707}_{3}$
(figure 11
c,d), the reference value about which variations occur is significantly increased.
4.5 Orientation, velocity and stream function
Finally, to understand how the fibre orientation and velocity are impacted by the anisotropic fluid properties, the orientation angle (
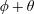 $\unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}$
) and velocity are considered in active and passive regimes, and the stream function is considered in all four regimes of interest (figures 12–14 respectively). Each variable is plotted for one wavelength of the sheet,
$\unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}$
) and velocity are considered in active and passive regimes, and the stream function is considered in all four regimes of interest (figures 12–14 respectively). Each variable is plotted for one wavelength of the sheet,
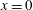 $x=0$
–
$x=0$
–
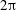 $2\unicode[STIX]{x03C0}$
. We focus on the case where the fibres are aligned with the sheet, i.e.
$2\unicode[STIX]{x03C0}$
. We focus on the case where the fibres are aligned with the sheet, i.e.
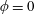 $\unicode[STIX]{x1D719}=0$
, and plot results at time
$\unicode[STIX]{x1D719}=0$
, and plot results at time
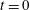 $t=0$
(i.e. the start of one oscillation period). See the supplementary movies available at https://doi.org/10.1017/jfm.2016.821.
$t=0$
(i.e. the start of one oscillation period). See the supplementary movies available at https://doi.org/10.1017/jfm.2016.821.
Considering first the fibre orientation, in all cases, perturbations to the initial orientation angle are greater in the vicinity of the sheet and are displaced with the movement of the sheet (figure 12 and movie 1). For passive rheology, the fibre reorientation is dampened very quickly moving away from the sheet (figure 12
a). Once
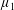 $\unicode[STIX]{x1D707}_{1}$
is non-zero, fibre displacement appears further away from the sheet (figure 12
b) and movements propagate to the right.
$\unicode[STIX]{x1D707}_{1}$
is non-zero, fibre displacement appears further away from the sheet (figure 12
b) and movements propagate to the right.
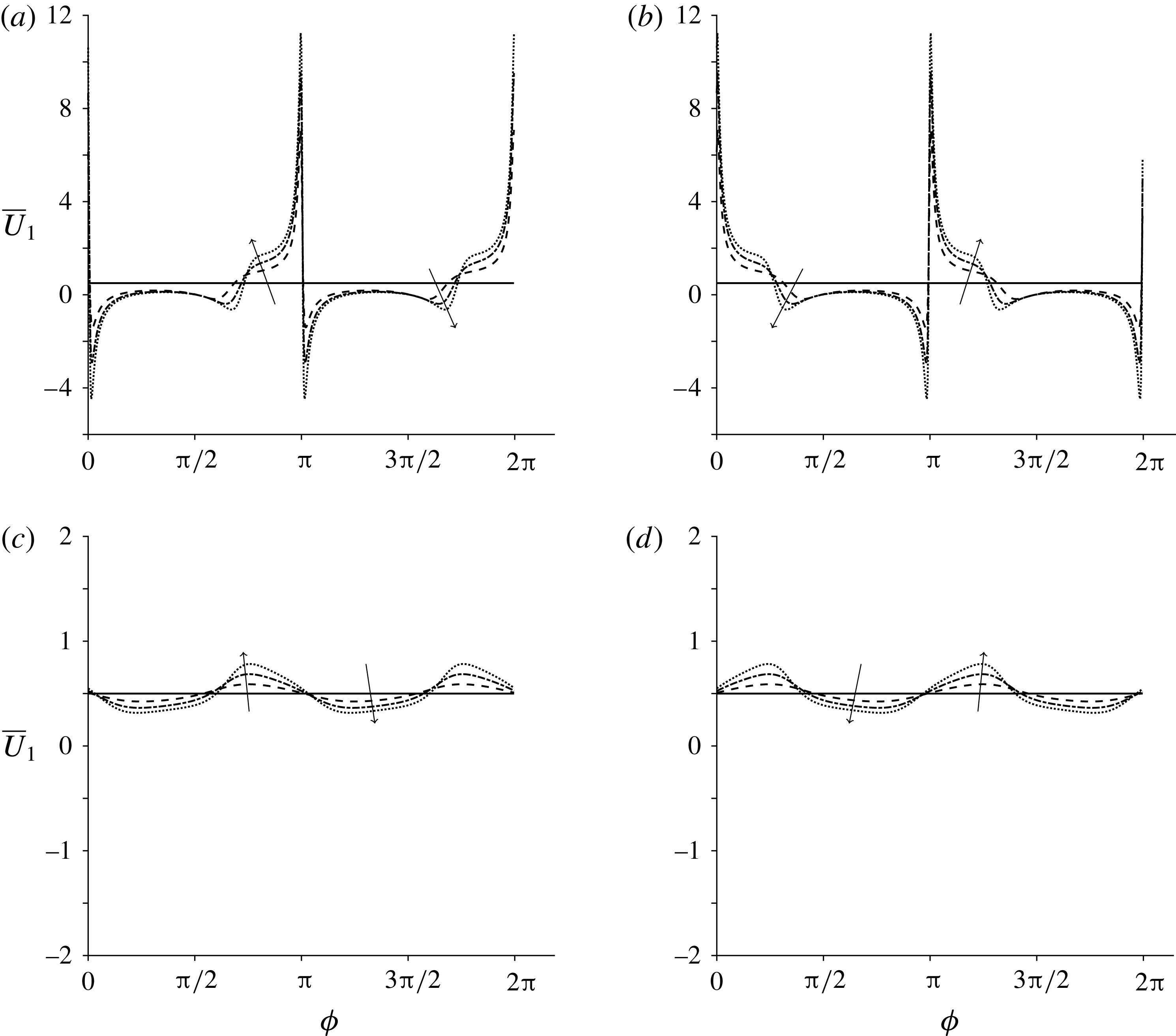
Figure 10. Mean swimming velocity versus
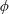 $\unicode[STIX]{x1D719}$
when at least one of
$\unicode[STIX]{x1D719}$
when at least one of
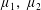 $\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{1},\,\unicode[STIX]{x1D707}_{2}$
and
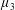 $\unicode[STIX]{x1D707}_{3}$
is much larger than 1. Panels (a) and (c) depict positive
$\unicode[STIX]{x1D707}_{3}$
is much larger than 1. Panels (a) and (c) depict positive
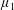 $\unicode[STIX]{x1D707}_{1}$
values and (b) and (d) depict negative
$\unicode[STIX]{x1D707}_{1}$
values and (b) and (d) depict negative
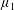 $\unicode[STIX]{x1D707}_{1}$
values. The values
$\unicode[STIX]{x1D707}_{1}$
values. The values
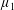 $\unicode[STIX]{x1D707}_{1}$
takes are
$\unicode[STIX]{x1D707}_{1}$
takes are
 $0$
(solid line),
$0$
(solid line),
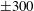 $\pm 300$
(dashed line),
$\pm 300$
(dashed line),
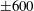 $\pm 600$
(dash-dotted line) and
$\pm 600$
(dash-dotted line) and
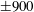 $\pm 900$
(dotted line). In (a) and (b),
$\pm 900$
(dotted line). In (a) and (b),
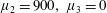 $\unicode[STIX]{x1D707}_{2}=900,\,\unicode[STIX]{x1D707}_{3}=0$
, and in (c) and (d),
$\unicode[STIX]{x1D707}_{2}=900,\,\unicode[STIX]{x1D707}_{3}=0$
, and in (c) and (d),
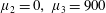 $\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=900$
, where the arrows denote increasing
$\unicode[STIX]{x1D707}_{2}=0,\,\unicode[STIX]{x1D707}_{3}=900$
, where the arrows denote increasing
 $\unicode[STIX]{x1D707}_{1}$
.
$\unicode[STIX]{x1D707}_{1}$
.
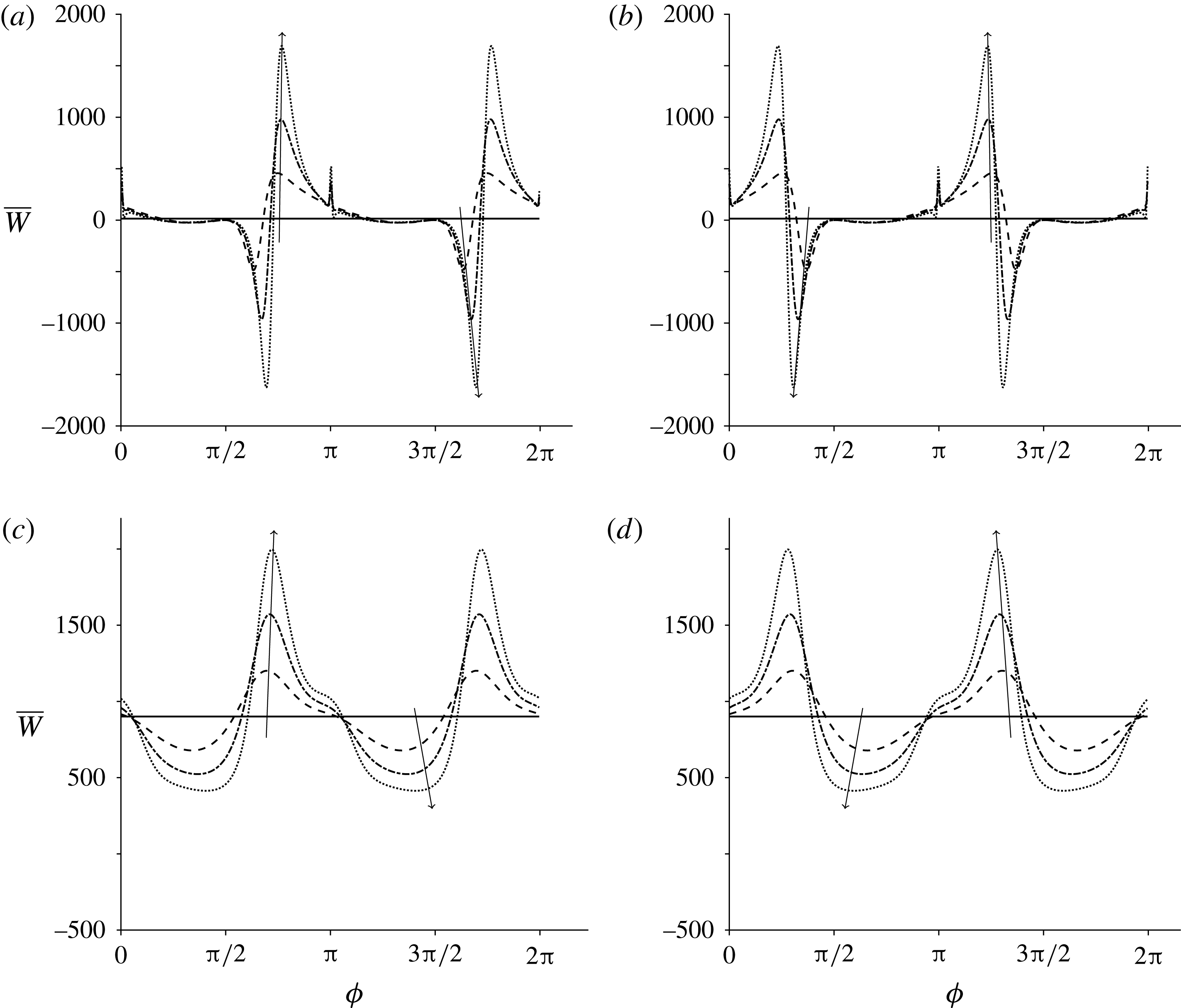
Figure 11. Mean rate of working versus
 $\unicode[STIX]{x1D719}$
when at least one of
$\unicode[STIX]{x1D719}$
when at least one of
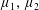 $\unicode[STIX]{x1D707}_{1},\unicode[STIX]{x1D707}_{2}$
and
$\unicode[STIX]{x1D707}_{1},\unicode[STIX]{x1D707}_{2}$
and
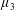 $\unicode[STIX]{x1D707}_{3}$
is much larger than 1. Panels (a) and (c) depict positive
$\unicode[STIX]{x1D707}_{3}$
is much larger than 1. Panels (a) and (c) depict positive
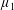 $\unicode[STIX]{x1D707}_{1}$
values and (b) and (d) depict negative
$\unicode[STIX]{x1D707}_{1}$
values and (b) and (d) depict negative
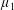 $\unicode[STIX]{x1D707}_{1}$
values. The values
$\unicode[STIX]{x1D707}_{1}$
values. The values
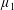 $\unicode[STIX]{x1D707}_{1}$
takes are
$\unicode[STIX]{x1D707}_{1}$
takes are
 $0$
(solid line),
$0$
(solid line),
 $\pm 300$
(dashed line),
$\pm 300$
(dashed line),
 $\pm 600$
(dash-dotted line) and
$\pm 600$
(dash-dotted line) and
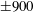 $\pm 900$
(dotted line). In (a) and (b),
$\pm 900$
(dotted line). In (a) and (b),
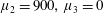 $\unicode[STIX]{x1D707}_{2}=900,\unicode[STIX]{x1D707}_{3}=0$
, and in (c) and (d),
$\unicode[STIX]{x1D707}_{2}=900,\unicode[STIX]{x1D707}_{3}=0$
, and in (c) and (d),
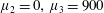 $\unicode[STIX]{x1D707}_{2}=0,\unicode[STIX]{x1D707}_{3}=900$
, where the arrows denote increasing
$\unicode[STIX]{x1D707}_{2}=0,\unicode[STIX]{x1D707}_{3}=900$
, where the arrows denote increasing
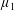 $\unicode[STIX]{x1D707}_{1}$
.
$\unicode[STIX]{x1D707}_{1}$
.

Figure 12. Fibre angle,
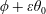 $\unicode[STIX]{x1D719}+\unicode[STIX]{x1D700}\unicode[STIX]{x1D703}_{0}$
, in passive and active regimes: (a) the passive regime (
$\unicode[STIX]{x1D719}+\unicode[STIX]{x1D700}\unicode[STIX]{x1D703}_{0}$
, in passive and active regimes: (a) the passive regime (
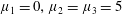 $\unicode[STIX]{x1D707}_{1}=0,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=5$
) and (b) the active-only regime (
$\unicode[STIX]{x1D707}_{1}=0,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=5$
) and (b) the active-only regime (
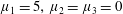 $\unicode[STIX]{x1D707}_{1}=5,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
). In each panel,
$\unicode[STIX]{x1D707}_{1}=5,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
). In each panel,
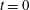 $t=0$
,
$t=0$
,
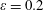 $\unicode[STIX]{x1D700}=0.2$
and the initial orientation angle is
$\unicode[STIX]{x1D700}=0.2$
and the initial orientation angle is
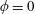 $\unicode[STIX]{x1D719}=0$
. See movie 1.
$\unicode[STIX]{x1D719}=0$
. See movie 1.
Figure 13 and movie 2 show the velocity components in
 $x$
and
$x$
and
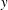 $y$
. The leading-order velocity decays quickly moving away from the sheet, as is evident in figure 13(a); in the active-only regime (figure 13
b), the flow decays more slowly. The velocity field shows a similar rightward propagation to fibre angle in the active-fluid case (figures 12
b and 13
b). These results are mirrored in figure 14 and movie 3, where the streamlines of the resulting flow are displayed. In the passive regime (i), the streamlines are symmetric about
$y$
. The leading-order velocity decays quickly moving away from the sheet, as is evident in figure 13(a); in the active-only regime (figure 13
b), the flow decays more slowly. The velocity field shows a similar rightward propagation to fibre angle in the active-fluid case (figures 12
b and 13
b). These results are mirrored in figure 14 and movie 3, where the streamlines of the resulting flow are displayed. In the passive regime (i), the streamlines are symmetric about
 $x=\unicode[STIX]{x03C0}$
with anticlockwise flow between
$x=\unicode[STIX]{x03C0}$
with anticlockwise flow between
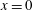 $x=0$
and
$x=0$
and
 $\unicode[STIX]{x03C0}$
and clockwise flow for
$\unicode[STIX]{x03C0}$
and clockwise flow for
 $x=\unicode[STIX]{x03C0}$
–
$x=\unicode[STIX]{x03C0}$
–
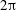 $2\unicode[STIX]{x03C0}$
. Introducing
$2\unicode[STIX]{x03C0}$
. Introducing
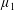 $\unicode[STIX]{x1D707}_{1}$
distorts the streamlines, and, when
$\unicode[STIX]{x1D707}_{1}$
distorts the streamlines, and, when
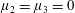 $\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
, the streamlines are deflected to the right (figure 14
b); introducing the other two parameters dampens this deflection (figure 14
c,d).
$\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
, the streamlines are deflected to the right (figure 14
b); introducing the other two parameters dampens this deflection (figure 14
c,d).
5 Discussion
The classical Taylor’s swimming sheet problem was modified to account for transverse isotropy, modelling swimming in fibre-reinforced fluids or active media. The quantities of interest were the steady background flow, which corresponds to the mean swimming velocity, and the mean rate of working. The results presented were non-dimensional. The dimensional velocity scales with the wave speed, and the rate of working scales with the square of the frequency, the viscosity and the wavenumber. The ratio of the mean swimming speed to the wave speed is proportional to
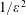 $1/\unicode[STIX]{x1D700}^{2}$
. When
$1/\unicode[STIX]{x1D700}^{2}$
. When
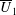 $\overline{U}_{1}$
takes the maximum value found here, such that
$\overline{U}_{1}$
takes the maximum value found here, such that
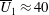 $\overline{U}_{1}\approx 40$
, this corresponds to swimming faster than the wave speed when
$\overline{U}_{1}\approx 40$
, this corresponds to swimming faster than the wave speed when
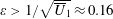 $\unicode[STIX]{x1D700}>1/\sqrt{\overline{U}_{1}}\approx 0.16$
. It should be noted that swimming with a prescribed beat amplitude and frequency, regardless of the rheology of the fluid, will not in general be achievable in a real biological system.
$\unicode[STIX]{x1D700}>1/\sqrt{\overline{U}_{1}}\approx 0.16$
. It should be noted that swimming with a prescribed beat amplitude and frequency, regardless of the rheology of the fluid, will not in general be achievable in a real biological system.

Figure 13. Velocity field in passive and active regimes: (a) passive regime (
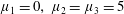 $\unicode[STIX]{x1D707}_{1}=0,\,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=5$
) and (b) active-only regime (
$\unicode[STIX]{x1D707}_{1}=0,\,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=5$
) and (b) active-only regime (
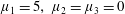 $\unicode[STIX]{x1D707}_{1}=5,\,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
). In each panel,
$\unicode[STIX]{x1D707}_{1}=5,\,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
). In each panel,
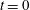 $t=0$
and the initial orientation angle is
$t=0$
and the initial orientation angle is
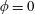 $\unicode[STIX]{x1D719}=0$
. See movie 2.
$\unicode[STIX]{x1D719}=0$
. See movie 2.

Figure 14. Instantaneous streamlines in the four regimes considered: (a) passive regime (
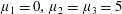 $\unicode[STIX]{x1D707}_{1}=0,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=5$
), (b) active-only regime (
$\unicode[STIX]{x1D707}_{1}=0,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=5$
), (b) active-only regime (
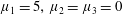 $\unicode[STIX]{x1D707}_{1}=5,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
), (c) nearly isotropic regime (
$\unicode[STIX]{x1D707}_{1}=5,\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
), (c) nearly isotropic regime (
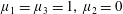 $\unicode[STIX]{x1D707}_{1}=\unicode[STIX]{x1D707}_{3}=1,\unicode[STIX]{x1D707}_{2}=0$
) and (d) regime where at least one of the parameters is much larger than 1 (
$\unicode[STIX]{x1D707}_{1}=\unicode[STIX]{x1D707}_{3}=1,\unicode[STIX]{x1D707}_{2}=0$
) and (d) regime where at least one of the parameters is much larger than 1 (
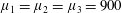 $\unicode[STIX]{x1D707}_{1}=\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=900$
). In each panel,
$\unicode[STIX]{x1D707}_{1}=\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=900$
). In each panel,
 $t=0$
and the initial orientation angle is
$t=0$
and the initial orientation angle is
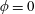 $\unicode[STIX]{x1D719}=0$
. See movie 3.
$\unicode[STIX]{x1D719}=0$
. See movie 3.
When
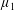 $\unicode[STIX]{x1D707}_{1}$
is zero, no dependence on the initial orientation angle is observed and the mean swimming velocity takes on the Newtonian value, regardless of the size of the other parameters. For non-zero
$\unicode[STIX]{x1D707}_{1}$
is zero, no dependence on the initial orientation angle is observed and the mean swimming velocity takes on the Newtonian value, regardless of the size of the other parameters. For non-zero
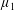 $\unicode[STIX]{x1D707}_{1}$
, certain initial orientation angles enable less energetically costly but slower swimming, with lower mean rates of working and swimming velocities. The initial angles maximising the mean swimming velocity and the mean rate of working are not in general the same. When active fibres are parallel to the swimming direction, swimming is both faster and more energetically costly compared with active fibres perpendicular to the swimming direction. The sheet utilises the surrounding environment to boost its velocity, a result seen in Leshansky (Reference Leshansky2009) for point-like obstacles and Chrispell, Fauci & Shelley (Reference Chrispell, Fauci and Shelley2013) for swimming in viscoelastic fluids near walls. However, these authors also predict an increased swimming efficiency, a result not seen here. A change from ‘pusher’ to ‘puller’ behaviour (equivalent to a change in sign of
$\unicode[STIX]{x1D707}_{1}$
, certain initial orientation angles enable less energetically costly but slower swimming, with lower mean rates of working and swimming velocities. The initial angles maximising the mean swimming velocity and the mean rate of working are not in general the same. When active fibres are parallel to the swimming direction, swimming is both faster and more energetically costly compared with active fibres perpendicular to the swimming direction. The sheet utilises the surrounding environment to boost its velocity, a result seen in Leshansky (Reference Leshansky2009) for point-like obstacles and Chrispell, Fauci & Shelley (Reference Chrispell, Fauci and Shelley2013) for swimming in viscoelastic fluids near walls. However, these authors also predict an increased swimming efficiency, a result not seen here. A change from ‘pusher’ to ‘puller’ behaviour (equivalent to a change in sign of
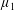 $\unicode[STIX]{x1D707}_{1}$
) equates to a reflection of the initial fibre angle in the
$\unicode[STIX]{x1D707}_{1}$
) equates to a reflection of the initial fibre angle in the
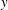 $y$
-axis. The activity parameter
$y$
-axis. The activity parameter
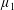 $\unicode[STIX]{x1D707}_{1}$
allows the mean rate of working to take on negative values, suggesting that the tension/stresslet exerted by the fibres on the sheet may at times overcome the work the sheet does on the fluid to move. For some values of
$\unicode[STIX]{x1D707}_{1}$
allows the mean rate of working to take on negative values, suggesting that the tension/stresslet exerted by the fibres on the sheet may at times overcome the work the sheet does on the fluid to move. For some values of
 $\unicode[STIX]{x1D707}_{1}$
, the mean swimming velocity may be negative, indicating a reversal of swimming direction; this change is dependent on the uniform orientation angle
$\unicode[STIX]{x1D707}_{1}$
, the mean swimming velocity may be negative, indicating a reversal of swimming direction; this change is dependent on the uniform orientation angle
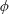 $\unicode[STIX]{x1D719}$
, a result also observed for rotated viscoelastic networks (Wróbel et al.
Reference Wróbel, Lynch, Barrett, Fauci and Cortez2016). The inclusion of active behaviour dramatically changes the streamlines and flow field.
$\unicode[STIX]{x1D719}$
, a result also observed for rotated viscoelastic networks (Wróbel et al.
Reference Wróbel, Lynch, Barrett, Fauci and Cortez2016). The inclusion of active behaviour dramatically changes the streamlines and flow field.
For a passive transversely isotropic fluid, i.e.
 $\unicode[STIX]{x1D707}_{1}=0$
, increasing the magnitude of the viscosity-like parameters increases the work the sheet must do on the fluid to swim; the fluid becomes more difficult to move through. The mean rate of working was found to be approximately linear in the parallel viscosity
$\unicode[STIX]{x1D707}_{1}=0$
, increasing the magnitude of the viscosity-like parameters increases the work the sheet must do on the fluid to swim; the fluid becomes more difficult to move through. The mean rate of working was found to be approximately linear in the parallel viscosity
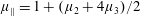 $\unicode[STIX]{x1D707}_{\Vert }=1+(\unicode[STIX]{x1D707}_{2}+4\unicode[STIX]{x1D707}_{3})/2$
, with a small additional dependence on
$\unicode[STIX]{x1D707}_{\Vert }=1+(\unicode[STIX]{x1D707}_{2}+4\unicode[STIX]{x1D707}_{3})/2$
, with a small additional dependence on
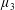 $\unicode[STIX]{x1D707}_{3}$
. For an active isotropic fluid (
$\unicode[STIX]{x1D707}_{3}$
. For an active isotropic fluid (
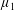 $\unicode[STIX]{x1D707}_{1}$
is non-zero and
$\unicode[STIX]{x1D707}_{1}$
is non-zero and
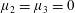 $\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
), we observe potentially unphysical behaviour when
$\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
), we observe potentially unphysical behaviour when
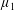 $\unicode[STIX]{x1D707}_{1}$
is increased sufficiently, with rapid large variations with respect to
$\unicode[STIX]{x1D707}_{1}$
is increased sufficiently, with rapid large variations with respect to
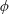 $\unicode[STIX]{x1D719}$
in both the mean swimming velocity and the rate of working. It should be noted, however, that a large value of
$\unicode[STIX]{x1D719}$
in both the mean swimming velocity and the rate of working. It should be noted, however, that a large value of
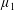 $\unicode[STIX]{x1D707}_{1}$
with
$\unicode[STIX]{x1D707}_{1}$
with
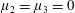 $\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
may not represent a physically realistic fluid. These unphysical effects are reduced by the inclusion of
$\unicode[STIX]{x1D707}_{2}=\unicode[STIX]{x1D707}_{3}=0$
may not represent a physically realistic fluid. These unphysical effects are reduced by the inclusion of
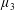 $\unicode[STIX]{x1D707}_{3}$
in particular, and to a lesser extent
$\unicode[STIX]{x1D707}_{3}$
in particular, and to a lesser extent
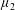 $\unicode[STIX]{x1D707}_{2}$
, dampening these large variations.
$\unicode[STIX]{x1D707}_{2}$
, dampening these large variations.
When the rheological parameters are all non-zero, increasing the anisotropic shear viscosity causes the mean swimming velocity to collapse down towards the Newtonian result, altered slightly by the active properties of the fluid. However, the mean rate of working is increased in general. The anisotropic extensional viscosity has a similar but much smaller effect.
This study demonstrates that locomotion in active fluids is dramatically different from locomotion in passive fluids. Our finding of zero, and indeed negative, rate of working for some angular configurations and sufficiently large values of the active parameter
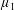 $\unicode[STIX]{x1D707}_{1}$
is a consequence of the energy input to the system by active stress. This phenomenon may be related to superfluid behaviour recently observed in bacteria suspensions (López et al.
Reference López, Gachelin, Douarche, Auradou and Clément2015). Further, these results are suggestive that active stresses in the bulk may enhance the motion of individual swimmers. It has long been known that some flagellated swimmers may propel more rapidly in polymeric than Newtonian fluids (Schneider & Doetsch Reference Schneider and Doetsch1974). An increase in the anisotropy of the slender-body drag coefficients has been proposed as one underlying mechanism (Berg & Turner Reference Berg and Turner1979; Magariyama & Kudo Reference Magariyama and Kudo2002); recently, Martinez et al. (Reference Martinez, Schwarz-Linek, Reufer, Wilson, Morozov and Poon2014) demonstrated that viscosity reduction associated with high-speed flagellar rotation provides an alternative explanation. The present model does not support a change to mean swimming speed purely through fluid anisotropy; because we analysed only 2D propulsion with constant viscosity-like parameters, we are unable to comment on the effect of shear-thinning on rotation.
$\unicode[STIX]{x1D707}_{1}$
is a consequence of the energy input to the system by active stress. This phenomenon may be related to superfluid behaviour recently observed in bacteria suspensions (López et al.
Reference López, Gachelin, Douarche, Auradou and Clément2015). Further, these results are suggestive that active stresses in the bulk may enhance the motion of individual swimmers. It has long been known that some flagellated swimmers may propel more rapidly in polymeric than Newtonian fluids (Schneider & Doetsch Reference Schneider and Doetsch1974). An increase in the anisotropy of the slender-body drag coefficients has been proposed as one underlying mechanism (Berg & Turner Reference Berg and Turner1979; Magariyama & Kudo Reference Magariyama and Kudo2002); recently, Martinez et al. (Reference Martinez, Schwarz-Linek, Reufer, Wilson, Morozov and Poon2014) demonstrated that viscosity reduction associated with high-speed flagellar rotation provides an alternative explanation. The present model does not support a change to mean swimming speed purely through fluid anisotropy; because we analysed only 2D propulsion with constant viscosity-like parameters, we are unable to comment on the effect of shear-thinning on rotation.
The passive region of parameter space with
 $\unicode[STIX]{x1D707}_{1}=0$
represents the anisotropic characteristics of the aligned passive microstructure of cervical mucus. Key aspects that may be explored in future work include shear-dependent viscosity and dispersion of fibre alignment. The active regime
$\unicode[STIX]{x1D707}_{1}=0$
represents the anisotropic characteristics of the aligned passive microstructure of cervical mucus. Key aspects that may be explored in future work include shear-dependent viscosity and dispersion of fibre alignment. The active regime
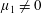 $\unicode[STIX]{x1D707}_{1}\neq 0$
may be considered as a model of motility through an active aligned medium, which may capture some of the essential physics of sperm migration through ciliated epithelium in the female reproductive tract. Our predictions could be tested experimentally by constructing an actuated membrane and studying the dynamics of an overlying suspension of swimming bacteria or microrods.
$\unicode[STIX]{x1D707}_{1}\neq 0$
may be considered as a model of motility through an active aligned medium, which may capture some of the essential physics of sperm migration through ciliated epithelium in the female reproductive tract. Our predictions could be tested experimentally by constructing an actuated membrane and studying the dynamics of an overlying suspension of swimming bacteria or microrods.
This study has opened up a number of exciting future research directions. These include (but are not limited to) investigating the effects of viscoelasticity (cf. Kruse et al., Reference Kruse, Joanny, Jülicher, Prost and Sekimoto2005), fibre dispersion (cf. Woodhouse & Goldstein, Reference Woodhouse and Goldstein2012) and the presence of walls (cf. Katz, Reference Katz1974). Similarly, coupling the flagellar elastic behaviour to the viscous fluid mechanics to determine the effect on the beat pattern (Riley & Lauga, Reference Riley and Lauga2014), and a full 3D computational treatment of the problem would be of interest. The model may also be developed to apply to peristaltic pumping by taking into account a cylindrical tube geometry. Taylor’s swimming sheet has inspired decades of research into biological propulsion; the study presented here shows that Taylor’s model continues to enable insight into novel areas of active fluid mechanics.
Acknowledgements
G.C. is supported by a Biotechnology and Biological Sciences Research Council (BBSRC) Industrial CASE Studentship (BB/L015587/1). The authors acknowledge C. Spalding and A. Tisbury for their contributions to an initial student project leading to this problem, and C. Holloway, Dr M. Gallagher and Dr M. Hicks for valuable discussions.
Appendix A. Components of the stress tensor
Assuming that the velocity takes the form
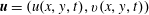 $\boldsymbol{u}=(u(x,y,t),v(x,y,t))$
, the components of the stress tensor are calculated as
$\boldsymbol{u}=(u(x,y,t),v(x,y,t))$
, the components of the stress tensor are calculated as
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D70E}_{11} & = & \displaystyle -p+\unicode[STIX]{x1D707}_{1}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}+ (\!2+\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{4}\nonumber\\ \displaystyle & & \displaystyle +\,4\unicode[STIX]{x1D707}_{3}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}\! ){\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}x^{}}}+ (\!\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{3}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\nonumber\\ \displaystyle & & \displaystyle +\,2\unicode[STIX]{x1D707}_{3}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\!)\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}y^{}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}y^{}}}\right)\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}y^{}}},\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D70E}_{11} & = & \displaystyle -p+\unicode[STIX]{x1D707}_{1}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}+ (\!2+\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{4}\nonumber\\ \displaystyle & & \displaystyle +\,4\unicode[STIX]{x1D707}_{3}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}\! ){\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}x^{}}}+ (\!\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{3}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\nonumber\\ \displaystyle & & \displaystyle +\,2\unicode[STIX]{x1D707}_{3}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\!)\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}y^{}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}y^{}}}\right)\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}y^{}}},\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D70E}_{12} & = & \displaystyle \unicode[STIX]{x1D70E}_{21}\nonumber\\ \displaystyle & = & \displaystyle \unicode[STIX]{x1D707}_{1}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\nonumber\\ \displaystyle & & \displaystyle +\,(1+\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}+\unicode[STIX]{x1D707}_{3}(1-\unicode[STIX]{x1D703}^{2}))\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}y^{}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}x^{}}}\right)\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{3}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719}){\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}x^{}}}\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{3}{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}y^{}}},\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D70E}_{12} & = & \displaystyle \unicode[STIX]{x1D70E}_{21}\nonumber\\ \displaystyle & = & \displaystyle \unicode[STIX]{x1D707}_{1}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\nonumber\\ \displaystyle & & \displaystyle +\,(1+\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}+\unicode[STIX]{x1D707}_{3}(1-\unicode[STIX]{x1D703}^{2}))\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}y^{}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}x^{}}}\right)\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{3}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719}){\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}x^{}}}\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{3}{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}y^{}}},\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D70E}_{22} & = & \displaystyle -p+\unicode[STIX]{x1D707}_{1}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}+ (\!2+\unicode[STIX]{x1D707}_{2}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{4}\nonumber\\ \displaystyle & & \displaystyle +\,4\unicode[STIX]{x1D707}_{3}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}\! ){\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}y^{}}}+ (\!\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{3}\nonumber\\ \displaystyle & & \displaystyle \times \,2\unicode[STIX]{x1D707}_{3}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\!)\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}y^{}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}x^{}}}\right)\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}{\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}x^{}}}.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D70E}_{22} & = & \displaystyle -p+\unicode[STIX]{x1D707}_{1}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}+ (\!2+\unicode[STIX]{x1D707}_{2}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{4}\nonumber\\ \displaystyle & & \displaystyle +\,4\unicode[STIX]{x1D707}_{3}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}\! ){\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}y^{}}}+ (\!\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{3}\nonumber\\ \displaystyle & & \displaystyle \times \,2\unicode[STIX]{x1D707}_{3}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})\!)\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}y^{}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{}v}{\unicode[STIX]{x2202}x^{}}}\right)\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}(\cos \unicode[STIX]{x1D719}-\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719})^{2}(\sin \unicode[STIX]{x1D719}+\unicode[STIX]{x1D703}\cos \unicode[STIX]{x1D719})^{2}{\displaystyle \frac{\unicode[STIX]{x2202}^{}u}{\unicode[STIX]{x2202}x^{}}}.\end{eqnarray}$$
Appendix B
The components of the matrix
 $\unicode[STIX]{x1D647}$
are calculated as
$\unicode[STIX]{x1D647}$
are calculated as
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D613}_{11} & = & \displaystyle \left(1+\frac{\unicode[STIX]{x1D707}_{2}}{4}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\right)(\unicode[STIX]{x1D706}^{4}-2\unicode[STIX]{x1D706}^{2}+1)-\unicode[STIX]{x1D707}_{2}\unicode[STIX]{x1D706}^{2}\cos 4\unicode[STIX]{x1D719}\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{1}[2\sin 2\unicode[STIX]{x1D719}(\unicode[STIX]{x1D706}\cos ^{2}\unicode[STIX]{x1D719}-\unicode[STIX]{x1D706}^{3}\sin ^{2}\unicode[STIX]{x1D719})+(\unicode[STIX]{x1D706}+\unicode[STIX]{x1D706}^{3})\cos 2\unicode[STIX]{x1D719}\sin 2\unicode[STIX]{x1D719}],\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D613}_{11} & = & \displaystyle \left(1+\frac{\unicode[STIX]{x1D707}_{2}}{4}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\right)(\unicode[STIX]{x1D706}^{4}-2\unicode[STIX]{x1D706}^{2}+1)-\unicode[STIX]{x1D707}_{2}\unicode[STIX]{x1D706}^{2}\cos 4\unicode[STIX]{x1D719}\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{1}[2\sin 2\unicode[STIX]{x1D719}(\unicode[STIX]{x1D706}\cos ^{2}\unicode[STIX]{x1D719}-\unicode[STIX]{x1D706}^{3}\sin ^{2}\unicode[STIX]{x1D719})+(\unicode[STIX]{x1D706}+\unicode[STIX]{x1D706}^{3})\cos 2\unicode[STIX]{x1D719}\sin 2\unicode[STIX]{x1D719}],\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D613}_{12} & = & \displaystyle -\unicode[STIX]{x1D707}_{1}[2\unicode[STIX]{x1D706}^{2}\sin ^{2}2\unicode[STIX]{x1D719}+\cos 2\unicode[STIX]{x1D719}((\unicode[STIX]{x1D706}^{2}+\unicode[STIX]{x1D706}^{4})\sin ^{2}\unicode[STIX]{x1D719}-(1+\unicode[STIX]{x1D706}^{2})\cos ^{2}\unicode[STIX]{x1D719})]\nonumber\\ \displaystyle & & \displaystyle +\,\frac{\unicode[STIX]{x1D707}_{2}}{2}(\unicode[STIX]{x1D706}^{3}+\unicode[STIX]{x1D706})\sin 4\unicode[STIX]{x1D719},\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D613}_{12} & = & \displaystyle -\unicode[STIX]{x1D707}_{1}[2\unicode[STIX]{x1D706}^{2}\sin ^{2}2\unicode[STIX]{x1D719}+\cos 2\unicode[STIX]{x1D719}((\unicode[STIX]{x1D706}^{2}+\unicode[STIX]{x1D706}^{4})\sin ^{2}\unicode[STIX]{x1D719}-(1+\unicode[STIX]{x1D706}^{2})\cos ^{2}\unicode[STIX]{x1D719})]\nonumber\\ \displaystyle & & \displaystyle +\,\frac{\unicode[STIX]{x1D707}_{2}}{2}(\unicode[STIX]{x1D706}^{3}+\unicode[STIX]{x1D706})\sin 4\unicode[STIX]{x1D719},\end{eqnarray}$$
where
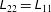 $\unicode[STIX]{x1D613}_{22}=\unicode[STIX]{x1D613}_{11}$
and
$\unicode[STIX]{x1D613}_{22}=\unicode[STIX]{x1D613}_{11}$
and
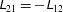 $\unicode[STIX]{x1D613}_{21}=-\unicode[STIX]{x1D613}_{12}$
.
$\unicode[STIX]{x1D613}_{21}=-\unicode[STIX]{x1D613}_{12}$
.
Appendix C
The balance of (3.3) at order
 $\unicode[STIX]{x1D700}^{2}$
is given by
$\unicode[STIX]{x1D700}^{2}$
is given by
 $$\begin{eqnarray}\displaystyle & & \displaystyle \left(1+\frac{\unicode[STIX]{x1D707}_{2}}{4}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\right)\unicode[STIX]{x1D6FB}^{4}\unicode[STIX]{x1D713}_{1}-\unicode[STIX]{x1D707}_{1}\left(2\sin 2\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{1}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+\cos 2\unicode[STIX]{x1D719}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{1}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{1}}{\unicode[STIX]{x2202}y^{2}}}\right)\right)\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}\left(\cos 4\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+\frac{\sin 4\unicode[STIX]{x1D719}}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}\right)\right)\nonumber\\ \displaystyle & & \displaystyle -\,\unicode[STIX]{x1D707}_{1}\left[2\sin 2\unicode[STIX]{x1D719}\left(\unicode[STIX]{x1D703}_{0}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{2}}}\right)+\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{}}}\right)^{2}-\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}}}\right)^{2}\right)\right.\nonumber\\ \displaystyle & & \displaystyle \left.+\,\cos 2\unicode[STIX]{x1D719}\left(4\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{}}}+\unicode[STIX]{x1D703}_{0}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right)\right)\right]-\unicode[STIX]{x1D707}_{2}\left[\sin 4\unicode[STIX]{x1D719}\left(2{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right.\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,\frac{\unicode[STIX]{x1D703}_{0}}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{4}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{4}}}\right)+\frac{1}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{2}}}\right)\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{3}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{}}}\right)+{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}}}\left(3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{3}}}\right)+\unicode[STIX]{x1D703}_{0}{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}\right)\right.\nonumber\\ \displaystyle & & \displaystyle \left.+\,\cos 4\unicode[STIX]{x1D719}\left(2\unicode[STIX]{x1D703}_{0}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}\right)+\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{2}}}\right){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right.\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{3}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{}}}\right)-{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{}}}\left(3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{3}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}}}\right)\right)\right]=0.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & & \displaystyle \left(1+\frac{\unicode[STIX]{x1D707}_{2}}{4}\sin ^{2}2\unicode[STIX]{x1D719}+\unicode[STIX]{x1D707}_{3}\right)\unicode[STIX]{x1D6FB}^{4}\unicode[STIX]{x1D713}_{1}-\unicode[STIX]{x1D707}_{1}\left(2\sin 2\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{1}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}+\cos 2\unicode[STIX]{x1D719}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{1}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{1}}{\unicode[STIX]{x2202}y^{2}}}\right)\right)\nonumber\\ \displaystyle & & \displaystyle +\,\unicode[STIX]{x1D707}_{2}\left(\cos 4\unicode[STIX]{x1D719}{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+\frac{\sin 4\unicode[STIX]{x1D719}}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{1}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}\right)\right)\nonumber\\ \displaystyle & & \displaystyle -\,\unicode[STIX]{x1D707}_{1}\left[2\sin 2\unicode[STIX]{x1D719}\left(\unicode[STIX]{x1D703}_{0}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{2}}}\right)+\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{}}}\right)^{2}-\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}}}\right)^{2}\right)\right.\nonumber\\ \displaystyle & & \displaystyle \left.+\,\cos 2\unicode[STIX]{x1D719}\left(4\left({\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{}}}+\unicode[STIX]{x1D703}_{0}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right)\right)\right]-\unicode[STIX]{x1D707}_{2}\left[\sin 4\unicode[STIX]{x1D719}\left(2{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right.\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,\frac{\unicode[STIX]{x1D703}_{0}}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{4}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}+{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{4}}}\right)+\frac{1}{2}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{2}}}\right)\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{3}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{}}}\right)+{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}}}\left(3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{3}}}\right)+\unicode[STIX]{x1D703}_{0}{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{2}}}\right)\right.\nonumber\\ \displaystyle & & \displaystyle \left.+\,\cos 4\unicode[STIX]{x1D719}\left(2\unicode[STIX]{x1D703}_{0}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{3}\unicode[STIX]{x2202}y^{}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{4}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{3}}}\right)+\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{2}}}\right){\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\right.\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{3}}}-3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}\unicode[STIX]{x2202}y^{}}}\right)-{\displaystyle \frac{\unicode[STIX]{x2202}^{}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}y^{}}}\left(3{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{3}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{3}}}\right)\right.\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D703}_{0}}{\unicode[STIX]{x2202}x^{}\unicode[STIX]{x2202}y^{}}}\left({\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}y^{2}}}-{\displaystyle \frac{\unicode[STIX]{x2202}^{2}\unicode[STIX]{x1D713}_{0}}{\unicode[STIX]{x2202}x^{2}}}\right)\right)\right]=0.\end{eqnarray}$$
Supplementary movies
Supplementary movies are available at https://doi.org/10.1017/jfm.2016.821.
















 Open access
Open access