


Introduction
In the European Union, animal welfare legislation (European Council, 2001) imposes the provision of bulky or high-fibre (HF) diets to pregnant sows to improve sow welfare. Such practices intend to fulfil feeding motivation in sows by increasing the daily feed allowance without increasing the energy intake during gestation, and thus avoiding overweight at farrowing which can be detrimental for piglets’ survival (Dourmad et al., Reference Dourmad, Etienne, Prunier and Noblet1994). Many studies have highlighted the beneficial effects of fibrous components in the gestation diet or through the litter on sow welfare (Meunier-Salaün et al., Reference Meunier-Salaün, Edwards and Robert2001). Dietary fibres are also known to reduce sow digestive disorders such as constipation (Wenk, Reference Wenk2001; Tabeling et al., Reference Tabeling, Schwier and Kamphues2003) as well as excessive body fatness at farrowing, that has been shown to lead to longer parturition length and increased piglet losses (Bilkei Papp, Reference Bilkei Papp1990). Approximately 75% of the 8% of stillborn piglets generally reported in usual rearing conditions die during parturition (Van Dijk et al., Reference Van Dijk, Van Rens, Van Der Lende and Taverne2005), mainly as a result of asphyxia (Van Rens and Van Der Lende, Reference Van Rens and Van Der Lende2004). Last-born piglets are more likely to suffer from asphyxia resulting from the cumulative effects of successive contractions or from a rupture of umbilical cord as parturition progresses (Randall, Reference Randall1971 and Reference Randall1972; Herpin et al., Reference Herpin, Le Dividich, Hulin, Fillaut, De Marco and Bertin1996). Live-born piglets experiencing asphyxia just before birth may suffer reduced early postnatal vitality which compromises their growth and survival rate until 10 days of age (Herpin et al., Reference Herpin, Le Dividich, Hulin, Fillaut, De Marco and Bertin1996). The death of live-born piglets can also be related to sow’s behaviour. It can result from low attention of the sow towards the piglets, which increases the risk of crushing (Grandinson et al., Reference Grandinson, Rydhmer, Strandberg and Thodberg2003), or from sow’s nervousness combined with aggressiveness and further infanticide. Infanticide can explain up to 25% of piglet losses (Ahlström et al., Reference Ahlström, Jarvis and Lawrence2002) and was reported especially in primiparous sows (Randall, Reference Randall1972). Although the few studies conducted on the effects of dietary fibre on parturition progress seem to show beneficial consequences of bulky diets through the reduction of parturition length, the effects on sow reproductive performance are less clear. Some authors report no effect of the addition of dietary fibre in the gestation diet on piglet growth (Courboulay and Gaudré, Reference Courboulay and Gaudré2002; Holt et al., Reference Holt, Johnston, Baidoo and Shurson2006), whereas others report either a positive (Matte et al., Reference Matte, Robert, Girard, Farmer and Martineau1994) or a negative (Vestergaard and Danielsen, Reference Vestergaard and Danielsen1998) effect of such a diet.
Thus, it clearly appears that parturition progress, in terms of timing of birth progress and sow behaviour, is of major relevance in peri-partum survival and subsequent performance of piglets. Many investigations have been carried out on the impact of animal factors, such as breed, parity, litter size, or piglet weight on parturition progress (Van Rens and Van Der Lende, Reference Van Rens and Van Der Lende2004; Van Dijk et al., Reference Van Dijk, Van Rens, Van Der Lende and Taverne2005). The scarce available data on the impact of environmental factors are mainly focused on pen design (Fraser et al., Reference Fraser, Phillips and Thompson1997) or on thermal conditions (Tantasuparuk et al., Reference Tantasuparuk, Lundeheim, Dalin, Kunavongkrit and Einarsson2000) rather than on dietary factors. Previous studies on the effect of supplying fibrous diets during gestation suggested that parturition duration was reduced by 9% to 29% according to the experiment. However, these studies differed in the breeds of sows used, sources of dietary fibre used and duration of allocation of the fibrous diet (Morgenthum and Bolduan, Reference Morgenthum and Bolduan1988; Kurcman-Przedpełska, Reference Kurcman-Przedpełska1989; Bilkei Papp, Reference Bilkei Papp1990). Thus, the effect of a fibrous diet on parturition progress and its impact on piglets’ performance needs to be confirmed. Therefore, the aim of the present study was to investigate the effect of a HF diet given throughout gestation on the parturition progress, and on sow and piglets performance.
Material and methods
Animals and housing
Forty-two primiparous sows (Large White × Landrace crossbed), averaging 143 ± 6 kg of live weight at mating, were used in two trials involving 24 and 18 sows, respectively (three replicates with eight and six sows per replicate in trials 1 and 2, respectively). In trial 1, the farrowing pen was equipped with a computerised system connected to a load cell located under the trough in order to record the individual feeding behaviour of sows during lactation (Guillemet et al., Reference Guillemet, Dourmad and Meunier-Salaün2006a); in trial 2, the sows were equipped with a carotid catheter for the measurement of metabolic and endocrine parameters around parturition and during lactation (unpublished data). Both trials were conducted following the same experimental design and housing conditions. During the first 5 weeks of gestation, all sows were individually housed in stalls measuring 1.90 × 0.60 m, on concrete floor with wood shavings. Thereafter sows were maintained in groups of four in pens measuring 4.20 × 2.30 m, with concrete floor and wood shavings, until the 15th week of gestation. Pens were divided into a resting and a feeding area, the latter being equipped with individual stalls and troughs. Twelve days before the expected time of parturition, sows were moved into farrowing crates in a parturition room where they stayed until weaning. Farrowing crates were equipped with slatted floor and measured 2.60 × 1.60 m. The area available for the sow measured 1.90 × 0.60 m. In each parturition room containing eight (trial 1) or six (trial 2) farrowing crates, half of the sows belonged to each experimental treatment.
During the whole experimental period, artificial lighting was provided from 0800 to 1800 h (103 lx) and lighting of lower intensity (10 lx) remained from 1800 to 0800 h to allow video recording. Average ambient temperatures were 20 ± 1°C (mean ± s.d.) during gestation and 22 ± 1°C during lactation. Two heat lamps were available in each farrowing crate, on the left and right sides of the sow, delivering a local temperature of 30 ± 1°C.
Litter size was standardised to 11 and 12 piglets within 48 h after parturition in trials 1 and 2, respectively. These litter sizes were chosen in accordance with litter size at birth and possibilities of piglet cross-fostering within dietary treatment group in each trial. On the 1st day post partum, piglets were ear-marked and received an iron injection. Males were castrated at 7 days of age. Weaning occurred during the 4th week post partum, on average 26.5 ± 0.4 days after parturition (mean ± s.d.).
Diets
From mating until the 4th week of gestation, all sows were fed once a day 2.50 kg of a standard gestation diet. From the 5th week of gestation until the day of farrowing, they were allocated to two experimental diets providing the same daily digestible energy (DE) supply (33.4 MJ) but containing either a low (control diet, C) or a high (high-fibre diet, HF) level of crude fibre (3.2% v. 12.4% of crude fibre, in % of dry matter (DM), respectively). The control diet was based on wheat, barley and soya-bean meal, whereas the wheat and soya-bean meal fractions were replaced by a mixture of fibre-rich feedstuffs of different types in the HF diet. Ingredient composition and chemical characteristics of both experimental diets are described in Table 1. Both diets were pelleted and provided once a day at 0900 h. Within each pen of four pregnant sows, half of the animals were individually fed the C diet and the HF diet (qualified as the C and HF sows, respectively). Diets were supplied after all sows of a group were locked in stalls where they were maintained during 75 min. At the end of the feeding period, sows were released in the resting area and feeding stalls were locked out. Any feed refusal was weighed if left in the trough. From the 1st day post partum until weaning, sows of both treatments were fed ad libitum the same standard lactation diet (Table 1). Water was available ad libitum in bowls during gestation and lactation. Piglets did not have any creep feed supply.
Table 1 Composition and chemical characteristics of the experimental diets
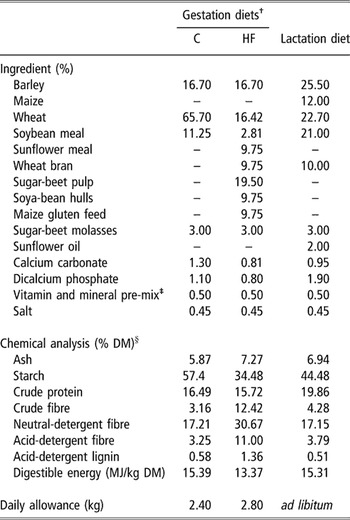
† C and HF, control diet and high-fibre diet during gestation, respectively.
‡ Contributed per kg, 10 000 IU vitamin A; 1500 IU vitamin D3; 30 mg vitamin E; 2 mg vitamin K3; 2 mg thiamine; 4 mg riboflavin; 20 mg niacin; 10 mg pantothenic acid; 3 mg pyridoxin; 0.02 mg biotin; 3 mg folic acid; 0.02 mg vitamin B12; 500 mg choline; 80 mg Fe; 10 mg Cu; 40 mg Mn; 100 mg Zn; 0.1 mg Co; 0.6 mg I; and 0.25 mg Se.
§ Percentage of dry matter (DM): 80%; digestible energy calculated from INRA-AFZ tables (2004).
Measurements
Parturition length and birth intervals
Parturition was not induced, and no medicine was administered during the whole parturition progress. For the 24 sows of trial 1, the parturition progress was videotaped without any intervention of stockpersons. In trial 2, the parturition progress was recorded de visu by an observer staying behind the sow during the whole duration of parturition. For all sows (n = 42), duration of parturition and time of birth of each piglet were recorded. Time when the first piglet was born was considered as the beginning of parturition. The duration of parturition was defined as the time between birth of the first and birth of the last piglet of the litter. For each piglet, the birth interval was defined as the time elapsed between two successive births. No medicine was administered during parturition.
Sow behaviour during parturition
Behavioural data were recorded on the 24 sows of trial 1 through the analysis of the continuous video-recordings. During parturition, the time spent by the sow in each posture and behavioural activities were measured. Postures were defined as lateral recumbency, ventral recumbency, sitting and standing, and thereafter the number of postural changes was determined. Behavioural activities included the following items: nesting behaviour (rooting, scratching and pawing at the pen floor), activities linked to uterine contractions, pen investigation, resting, piglet-directed social behaviour (visual attention towards a piglet with or without any physical contact), aggression (biting, pushing) and crushing. The sow was considered having uterine contractions when contractions of the rear part of the body and movements of the upper hind leg were detected.
Reproductive performance
The two trials were designed to study the effect of fibrous diet on reproductive performance. For all experimental sows, each piglet born was recorded as live-born, stillborn or mummified. However, due to difficulties encountered in identifying mummified piglets on video-recordings, these piglets were not included in the analysis. Piglets were weighed within 24 h of birth (day 0), on days 7, 14, 21 and at weaning.
Sow weight and backfat thickness were measured at mating, on day 105 of gestation, on the 1st day post partum, and at weaning. Backfat thickness was measured by ultrasonography (Sonolayer SAL-32B, Toshiba, Tokyo, Japan) at 65 mm on each side of the dorsal mid line at the last rib level (P2). Sow’s daily feed intake during lactation was measured each morning by weighing daily feed refusals.
Gestation length was defined as the interval between the day of the first artificial insemination and the day of parturition. Oestrus after weaning was detected by daily boar exposure and was only measured in the first trial.
Statistical analysis
Data on the birth progress were analysed for the 42 sows. Because data were not available in the second trial, the sow behaviour during parturition and the weaning-to-oestrus interval were analysed for the 24 sows of the first trial only. Sow and piglet performance were analysed for 41 sows: one sow from the control group had to be removed from the statistical analysis because of a prolonged period of fever that started after parturition.
Criteria measured once were analysed using the GLM procedure of Statistical Analysis Systems Institute (SAS, 2000), whereas criteria measured several times in the same sow, including birth intervals, body weight and backfat thickness of sows and weight of piglets, were analysed using a ‘repeated’ model with the MIXED procedure of SAS. For the latter criteria, sow within diet was considered as residual error in the analyses. For sow behavioural data, the time spent in each behavioural activity and posture was analysed after an arcsine transformation. Models tested the main effects of gestation diet, replicate and their interactions. Trial effect was also tested in the analysis of parameters related to the parturition progress and reproductive performance. Duration of parturition and piglet growth rate was adjusted for litter size and number of piglets weaned, respectively. Whatever the considered criteria, the trial × diet interaction was never significant. Results are expressed as means and standard errors of the mean (s.e.).
Results
Parturition and reproductive performance
Duration of gestation did not differ between the dietary treatments and averaged 115.2 ± 1.5 days (Table 2). The average duration of parturition was not significantly affected by the dietary treatments and amounted to 211 ± 77 min (Table 2). Sixty-nine per cent of all recorded parturitions were achieved within less than 4 h, 21% within 4 to 5 h, and 10% lasted more than 5 h. The longest parturition recorded lasted 403 min for 16 piglets, whereas the shortest parturition lasted 58 min for 13 piglets. In both cases, sows belonged to the HF treatment. Whatever the experimental treatment, litter size tended to affect the duration of parturition, larger litters needing more time to be delivered (P = 0.06). Our data followed the model:
where y is the duration of parturition (min), and x number of piglets born in the litter (mummified piglets are not included).
Table 2 Effects of trial (T), replicate (R) and gestation diet (D) on parturition and reproductive performance (means and residual standard deviation (r.s.d.), n = 42)

† C and HF, control diet (3.2% dry matter crude fibre) and high-fibre diet (12.4% DM crude fibre) during gestation, respectively (n = 21 sows per treatment).
‡ NS P > 0.10, *P < 0.05, **P < 0.01; the interaction T × R × D was significant on the birth interval P < 0.05.
§ Litter size at birth included the live born and the stillborn piglets.
∥ Criteria for the 24 sows of the first trial (n = 12 sows per treatment).
¶ Trial effect and interaction with other factors.
Birth interval was not affected by the dietary treatment (Table 2). Intervals averaged 16.5 ± 23 min but ranged from 0 to 242 min. Within this variation, 87% of the piglets were born less than 30 min after a littermate (Figure 1). Birth intervals were weakly affected by litter size (R 2=0.028).

Figure 1 Frequency of birth intervals between two successive littermates in all parturitions.
Birth interval between the first and the third born piglets was shorter in the HF sows than in the C sows (52 ± 12 and 40 ± 7 min, respectively) but this difference was not significant (Table 2). Weaning-to-oestrus interval did not differ between treatments and averaged 4.7 ± 0.5 days (Table 2). Significant differences were shown on the weaning-to-oestrus interval according to replicate, but without any clear pattern in values. In both trials, a total of 553 piglets were born among which 27 were stillborn (5%). The dietary treatment did not affect litter size at birth (in average 13.7 ± 3.1 piglets), percentage of stillborn piglets (in average 5.3 ± 7.7%) and number of piglets weaned (in average 11.2 ± 1.3 piglets) (Table 5). A trial effect on the percentage of stillborn piglets values was found (7.4 ± 1.5 v. 2.3 ± 0.8% in trials 1 and 2, respectively; P < 0.05) and on the number of weaned piglets (11.0 ± 0.2 v. 11.5 ± 0.2 weaned piglets in trials 1 and 2, respectively; P < 0.05). In all cases, the trial × diet interaction was never significant.
The dietary treatments did not have any significant effect on the behavioural criteria measured during the parturition of the 24 sows from the first trial (Table 3). Sows spent most of the time exhibiting uterine contractions (68 ± 24% of the time in average for both dietary treatments). The remainder of the time was mainly devoted to social behaviours with piglets, nesting behaviour and resting (in average 25 ± 20.5 and 2 ± 9%, respectively). Piglet-directed aggressiveness was observed in two sows only and occurred seven and four times in one C sow and in one HF sow, respectively. Time spent in the different postures did not differ between dietary treatments. The major posture adopted by the sows was lateral recumbency (62 ± 26% of the time in average, Table 3). Sows of both treatments spent the same amount of time standing (7 ± 7%) and sitting (22 ± 16%) and the number of postural changes did not differ significantly between treatments (in average 12 ± 8 changes). Some differences were found in criteria according to replicates, but without a clear pattern in values. For all criteria, no replicate × diet interaction was shown.
Table 3 Effects of replicate (R) and gestation diet (D) on the distribution of activities and postures in C sows and HF sows during parturition (means and residual standard deviation (r.s.d.), n = 24)

† C and HF, control diet (3·2% dry matter crude fibre) and high-fibre diet (12·4% dry matter crude fibre) during gestation, respectively (n = 12 sows per treatment).
‡ NS P > 0.10, *P < 0.05, **P < 0.01.
Changes in body weight of sows and piglets
The average daily feed intake (ADFI) of sows measured during the 23 first days of lactation was not affected by the dietary treatments and averaged 6.4 ± 1.2 kg/day (Table 4). Dietary treatments did not affect the variations of sows’ weight during gestation and lactation (Table 4). The HF sows gained less backfat thickness than the C sows during gestation (1.6 mm of difference, P < 0.05), but lost the same amount of backfat during lactation, leading to leaner HF sows at the end of pregnancy and at weaning (P < 0.05 and P < 0.01, respectively) (Table 4). The body weight of sows differed between trials at parturition and weaning, with greater values in trial 1 than in trial 2 (202 ± 2 v. 194 ± 3 kg, P < 0.01; 188 ± 3 v. 178 ± 4 kg, P < 0.05, respectively), but without any effect on the weight gain during gestation or the weight loss during lactation (Table 4). Limited significant differences were obtained according to trial or replicate, without a clear pattern in values. For all criteria, no interaction between factors was shown.
Table 4 Effects of trial (T), replicate (R) and gestation diet (D) on sow body weight and backfat thickness during gestation and lactation (means and residual standard deviation (r.s.d.), n = 42)
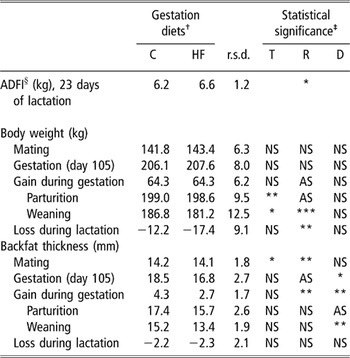
† C and HF, control diet (3·2% dry matter crude fibre) and high-fibre diet (12·4% dry matter crude fibre) during gestation, respectively (n = 12 sows per treatment).
‡ No significant interaction between factors T, R and D; significance level, NS P > 0.10, AS Approaching significance P < 0.10, *P < 0 .05, **P < 0.01.
§ Average daily feed intake.
Piglet weight at birth was similar in both treatments and averaged 1.40 ± 0.26 kg. During the 1st week of life, the piglets nursed by the HF sows had a 13.5% greater average daily weight gain compared with the piglets of the C sows (P < 0.01), but this difference was reduced over the following weeks (Table 5). Nevertheless, the impact of the 1st week induced heavier weights for the HF piglets on day 14 (P < 0.05) that tended to be maintained until day 21 and weaning (P = 0.06). The body weight of piglets measured weekly was lower in the second trial compared to the first one, whatever the considered period (P < 0.05, Table 5). Moreover, no trial × diet interaction was found on the growth rate variation of piglets. The daily weight gain differed between replicates during the week 4 only (P < 0.05).
Table 5 Effects of trial (T), replicate (R) and gestation diet (D) on growth rate of piglet (means and residual standard deviation, n = 42)
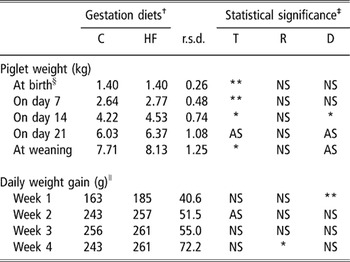
† C and HF, control diet (3.2% dry matter crude fibre) and high-fibre diet (12.4% dry matter crude fibre) during gestation, respectively (n = 21 sows/treatment).
‡ No significant interaction between factors T, R and D; significance level, NS P > 0.10, AS Approaching significance P < 0.10, *P < 0.05, **P < 0.01.
§ Litter size at birth included the live born and the stillborn piglets.
∥ Available for 20 and 21 sows of the C and HF diets, respectively.
Discussion
Parturition and reproductive performance
Our results did not show any significant effect of the dietary treatments on gestation length. Data available on the relationship between ingestion of a fibrous diet during gestation and gestation length in sows are very scarce. However, present results did not confirm the findings of Kurcman-Przedpełska Reference Kurcman-Przedpełska(1989) who reported a longer gestation length in sows fed a standard gestation diet compared with sows fed a fibrous diet. But in that study, the authors had used a different sow breed (Polish breed) and their experimental diets differed from ours by both their composition and their crude fibre content. An inverse relationship between gestation length and duration of parturition has been found by Van Rens and Van Der Lende Reference Van Rens and Van Der Lende(2004) who reported that an increased gestation length was associated with a shorter duration of parturition. In the present study, duration of parturition was not affected by dietary treatments and varied considerably between sows, ranging from 58 to 403 min. Compared with results reported in previous studies on primiparous sows, our values were either greater (on average 130 min, Van Rens and Van Der Lende, Reference Van Rens and Van Der Lende2004) or similar (on average 220 min, Bilkei Papp, Reference Bilkei Papp1990), but the discrepancy between studies could result from differences in litter sizes, breed, etc. In studies investigating the impact of dietary fibre supplied during gestation on the duration of parturition, it has been shown that sows fed a HF diet during gestation had shorter parturition durations than sows fed a standard gestation diet. The reduction was more or less significant and ranged from 9% to 29% according to authors (Morgenthum and Bolduan, Reference Morgenthum and Bolduan1988; Kurcman-Przedpełska, Reference Kurcman-Przedpełska1989; Bilkei Papp, Reference Bilkei Papp1990). Besides, Thodberg et al. Reference Thodberg, Jensen and Herskin(2002) suggested that a greater nervousness of the parturient sow could be associated to a longer duration of parturition but also to a shorter birth interval between the first- and the third-born piglets. As a consequence of the alleviating effects of a fibrous diet shown by Brouns et al. Reference Brouns, Edwards and English(1994) on the sow activity, it could be hypothesised that sows fed a fibrous diet during gestation would show less nervousness and consequently have shorter parturition durations. Nevertheless, we did not find such significant effect on the birth interval between the first and the third-born piglets and on the duration of parturition, which could be related to high variation between experimental sows for these criteria (min.–max.: 1–255 min, 58–403 min, respectively).
Van Rens and Van Der Lende Reference Van Rens and Van Der Lende(2004), as well as Van Dijk et al. Reference Van Dijk, Van Rens, Van Der Lende and Taverne(2005), found that an increased litter size resulted in a significantly lengthened duration of parturition. A same tendency was found in our study. A significant level might have not been achieved with our data because of an elevated mean litter (>12). Indeed, previous studies have reported no or a small relationship between litter size and parturition length when litter size was large (Friend et al., Reference Friend, Cunningham and Nicholson1962; Morgenthum and Bolduan, Reference Morgenthum and Bolduan1988).
Literature on duration of parturition appear somewhat confusing due to various definitions of this criterion among authors. Indeed, duration of parturition is either defined as the time interval between births of the first and the last piglet (Fahmy and Flipot, Reference Fahmy and Flipot1981; Van Rens and Van Der Lende, Reference Van Rens and Van Der Lende2004; Van Dijk et al., Reference Van Dijk, Van Rens, Van Der Lende and Taverne2005), or as the time interval between the birth of the first piglet and the expulsion of the placenta (Morgenthum and Bolduan, Reference Morgenthum and Bolduan1988). To evaluate more precisely the timing of the parturition progress, it would seem necessary to consider the birth intervals between two successive littermates rather than the mean time interval between the first piglet and the last one or the placenta expulsion. In the present study the mean birth interval was 16.5 ± 23 min, which is in agreement with the values of 16 and 15 min found by Randall Reference Randall(1972) and Van Rens and Van Der Lende Reference Van Rens and Van Der Lende(2004), respectively. This interval was not affected by the sow’s gestation diet. Morgenthum and Bolduan Reference Morgenthum and Bolduan(1988) indicated that a birth interval longer than 30 min increases the risk of hypoxia for the foetus, especially for the lightest and the last born ones. They also reported an average decrease from 31 to 26 min for the interval between successive births in sows fed a HF diet compared with sows fed a standard diet, and a number of stillborn piglets reduced by 16% in sows fed the fibrous diet. In our study, the birth intervals were similar and in average, 87% of them lasted less than 30 min in both treatments, which can explain the lack of a dietary effect on the rate of stillborn piglets. Litter size did not differ between dietary treatments in the present study, in agreement with the results of Holt et al. Reference Holt, Johnston, Baidoo and Shurson(2006).
Our results did not show any alleviating effect of the supply of dietary fibre in the gestation diet on sow behaviour during parturition, which could partly result from the important individual variation recorded on this criterion.
Sow’s restlessness level has been studied through the frequency of postural changes during parturition (Jarvis et al., Reference Jarvis, McLean, Calvert, Deans, Chirnside and Lawrence1999; Ahlström et al., Reference Ahlström, Jarvis and Lawrence2002; Thodberg et al., Reference Thodberg, Jensen and Herskin2002). A greater incidence of piglet crushing is recorded when postural changes are frequent (Weary et al., Reference Weary, Pajor, Fraser and Honkanen1996). Postural changes were less frequent in the HF sows that appeared quieter than the C sows. Whatever the dietary treatment, sows were mainly in a lateral recumbency posture, as usually exhibited during parturition (Vestergaad and Hansen, Reference Vestergaad and Hansen1984). As well as promoting the social bonding between the sow and her piglets (Petersen et al., Reference Petersen, Recén and Vestergaard1990), such posture facilitates the access to the udder as a source of nutrition and warmth (Fraser, Reference Fraser1984).
During parturition, the sows’ piglet-directed behaviours appeared scarce, with no difference between dietary treatments. This low level has been shown to play in favour of piglets’ survival. Jarvis et al. Reference Jarvis, McLean, Calvert, Deans, Chirnside and Lawrence(1999) reported that farrowing sows are generally unresponsive to their piglets during parturition, and suggested that such unresponsiveness could be related to good maternal abilities. Similarly, Ahlström et al. Reference Ahlström, Jarvis and Lawrence(2002) showed that the sows that are more responsive towards their piglets through an important level of piglet investigation are more likely to become savaging sows. These authors also suggested that a low responsiveness of the sow may not necessarily reflect poor maternal ability, but may be related to the individual’s inability to cope with the farrowing situation. This inability might be more marked in primiparous sows.
Growth rate variation of sows and piglets
Variations of sows’ weight did not differ according to the dietary treatment, whatever the stage considered between mating and weaning. However, although the daily allowance of each diet was intended to provide the same amount of DE to all sows, sows fed the standard diet gained more backfat thickness during gestation than sows fed the HF diet. The difference in backfat thickness gain could be related to the level of net energy (NE) intake in the two groups of sows. Indeed, both diets provided 33.4 MJ DE per day to each sow, but the daily NE supply was lower for the sows fed the HF diet than for those fed the control diet, due to a lower efficiency of utilisation of DE from crude fibre than from starch. Besides, the adaptation period to the experimental diets was longer with the HF diet, resulting in a slightly lower mean daily feed consumption during gestation than expected in sows fed that diet (2.73 instead of 2.80 kg/day). Consequently, the lower daily consumption of NE by the HF sows (20.5 v. 22.8 MJ NE per day in C sows) could also explain the smaller backfat thickness gain of the HF sows during gestation. The difference in backfat thickness at farrowing between the HF and the C sows was maintained until weaning as all sows lost on average the same amount of backfat during lactation. Differences in backfat thickness at weaning did not influence the subsequent weaning-to-oestrus interval. The lack of effect of dietary fibre on the weaning-to-oestrus interval is consistent with the previous results reported in literature (Matte et al., Reference Matte, Robert, Girard, Farmer and Martineau1994; Farmer et al., Reference Farmer, Robert and Matte1996; Vestergaard and Danielsen, Reference Vestergaard and Danielsen1998; Holt et al., Reference Holt, Johnston, Baidoo and Shurson2006). Some differences were found between trials or replicates in backfat thickness without any clear pattern to explain theses effects. Various factors could be involved, such as the ambient temperature which is known to affect energy metabolism of pregnant sows (Noblet et al., Reference Noblet, Dourmad, Le Dividich and Dubois1989).
Variations of sows’ weight did not differ between the dietary treatments, whatever the stages considered between mating and weaning. The lack of impact on body weight could be explained by an increased weight of digestive tract in sows fed the HF diet during gestation. Such an impact on the digestive tract has been shown in growing pigs (Pond et al., Reference Pond, Jung and Varel1988; Jørgensen et al., Reference Jørgensen, Zhao and Eggum1996).
The weight of piglets at birth and their growth rate did not differ between the dietary treatments in the present study, which is in agreement with the results of Holt et al. Reference Holt, Johnston, Baidoo and Shurson(2006) using similar diets at iso-energetic levels (2.7 v. 14.8% of CF in the diet). However, Matte et al. Reference Matte, Robert, Girard, Farmer and Martineau(1994) found that during their second parity, sows fed a gestation diet based on wheat bran and maize cobs gave birth to heavier piglets than sows fed diets based on maize and soya-bean meal, or on oat hulls and oats. These piglets were also heavier at weaning. In our study, piglets issued from sows fed the HF diet showed an improved weight gain during the 1st week of life, were heavier (+7%) at mid lactation, and tended to be heavier at weaning (+5.5%). This improved growth rate might have a positive impact on further piglet survival, especially in highly prolific sows which nurse large litters (Rydhmer, Reference Rydhmer1992). Vestergaard and Danielsen Reference Vestergaard and Danielsen(1998) reported that piglets born from sows fed a low fibre diet (5.3% DM of crude fibre) were heavier at birth than piglets from sows fed a bulky diet based on sugar-beet pulp (13.2% DM of crude fibre), and this difference had disappeared at weaning. Such discrepancies between studies could be explained by numerous factors. Indeed, the impact of dietary fibre on sow reproductive performance widely differs according to sow parity, timing of fibre allowance, and source of fibrous feed ingredient included in the gestation diet, as reported by Holt et al. Reference Holt, Johnston, Baidoo and Shurson(2006). Because no creep feed was available, the faster growth of the HF piglets very likely resulted from greater milk and/or nutrient production from the sows fed the HF diet. This could have been the result of a more important piglet stimulation of the udder. An improved milk yield might have been made possible through the greater feed intake measured in HF sows. Although the feed intake was not significantly greater in these sows during the whole lactation, it was significantly greater during the 1st week of lactation (Guillemet et al., Reference Guillemet, Dourmad and Meunier-Salaün2006a and Reference Guillemet, Hamard, Quesnel, Père, Etienne, Dourmad and Meunier-Salaün2006b). Further investigations are needed to test these hypotheses. Comparison of the present data with the previous results remains difficult because of the various factors involved in the dietary treatment that was applied, especially the level of fibre inclusion and the origin of fibrous components. In addition, in many studies sows were restrictively fed during the first days after parturition, which was not the case in our study.
Conclusion
This study provides information on the impact of the incorporation of dietary fibre in the gestation diet on the parturition progress and its consequences on piglet performances. Feeding the HF diet during gestation had limited effects on the parturition progress and on reproductive performance, whereas it appeared to improve piglets’ growth during their 1st week of life, leading to live weights that tended to be heavier at weaning. Further investigations are needed to confirm the effects of such diets on the peri-partum period.
Acknowledgements
We would like to thank animal keepers and all the staff at the Research Centre who participated in the surveillance of the parturitions by night and day, especially Michel Lefebvre, Hervé Demay, Loïc Gaillard, Henri Renoult and Michel Massard. We also thank Pascal Bodinier and Morgane Moreau for the technical work, in particular for the analysis of video-recordings.






