



In northeastern North America (NENA), archaeological studies of agriculture have focused almost exclusively on when Indigenous people adopted maize and how much maize they cultivated (e.g., Chilton Reference Chilton and Hart2008; Hart Reference Hart2000, Reference Hart and Hart2008; Hart and Lovis Reference Hart and Lovis2013; Hart et al. Reference Hart, Brumbach and Lusteck2007; Katzenberg Reference Katzenberg, Staller, Tykot and Benz2006; cf., Asch Sidell Reference Asch Sidell and Hart2008; Messner Reference Messner2008). Despite the emphasis on maize, archaeologists have recognized that Indigenous people in the region also farmed beans (Phaseolus vulgaris), squash (Cucurbita pepo), and sunflower (Helianthus annuus var. macrocarpus and Helianthus tuberosus; e.g., Asch Sidell Reference Asch Sidell and Hart2008; Hart and Lovis Reference Hart and Lovis2013). Squash and sunflower were initially domesticated by Indigenous people farther to the west as part of what is now referred to as the Eastern Agricultural Complex (EAC).
Consisting of domesticated chenopodium (Chenopodium berlandieri ssp. jonesianum), marshelder (Iva annua var. macrocarpa), sunflower, squash, and erect knotweed (Polygonum erectum ssp. watsoniae), along with nondomesticated but cultivated plants including maygrass (Phalaris caroliniana) and little barley (Hordeum pusillum), the EAC formed an important component of Indigenous foodways west of the Appalachian Mountains for over 4,000 years (Fritz Reference Fritz2019; Mueller Reference Mueller2017, Reference Mueller2018; Mueller et al. Reference Mueller, Fritz, Patton, Carmody and Horton2017; Smith Reference Smith and Keegan1987, Reference Smith2007, Reference Smith2011).
Despite the dietary and culinary importance of EAC cultigens in broader eastern North America (ENA), paleoethnobotanists have infrequently documented EAC domesticates and cultivated species in NENA (Figure 1; Table 1; Asch and Hart Reference Asch, Hart and Goodman2004; Asch Sidell Reference Asch Sidell and Hart2008:Table 3-3; Crawford et al. Reference Crawford, Lytle, Williamson and Wojtowicz2019; Fritz Reference Fritz2019; Hart and Lovis Reference Hart and Lovis2013; Mueller et al. Reference Mueller, Fritz, Patton, Carmody and Horton2017; Scarry Reference Scarry and Minnis2003, Reference Scarry, Reitz, Scarry and Scudder2008; Smith Reference Smith2007). When encountered in NENA, EAC cultigens are usually found in small quantities and generally not in their domesticated forms (Asch Sidell Reference Asch Sidell and Hart2008:Table 3-3). Aside from squash and sunflower, which are more commonly identified as part of Late Woodland (ca. AD 800/900–contact) maize-agriculture systems in Ontario and New York, Indigenous people in NENA are generally not considered to have cultivated EAC plants (Asch Sidell Reference Asch Sidell and Hart2008; Crawford and Smith Reference Crawford, Smith and Minnis2003; Hart and Lovis Reference Hart and Lovis2013). Combined with the absence of mound building and the dearth of Hopewellian, Mississippian, or Fort Ancient–style artifacts, the lack of EAC cultigens in NENA has further separated the region as an outlier from the rest of ENA.
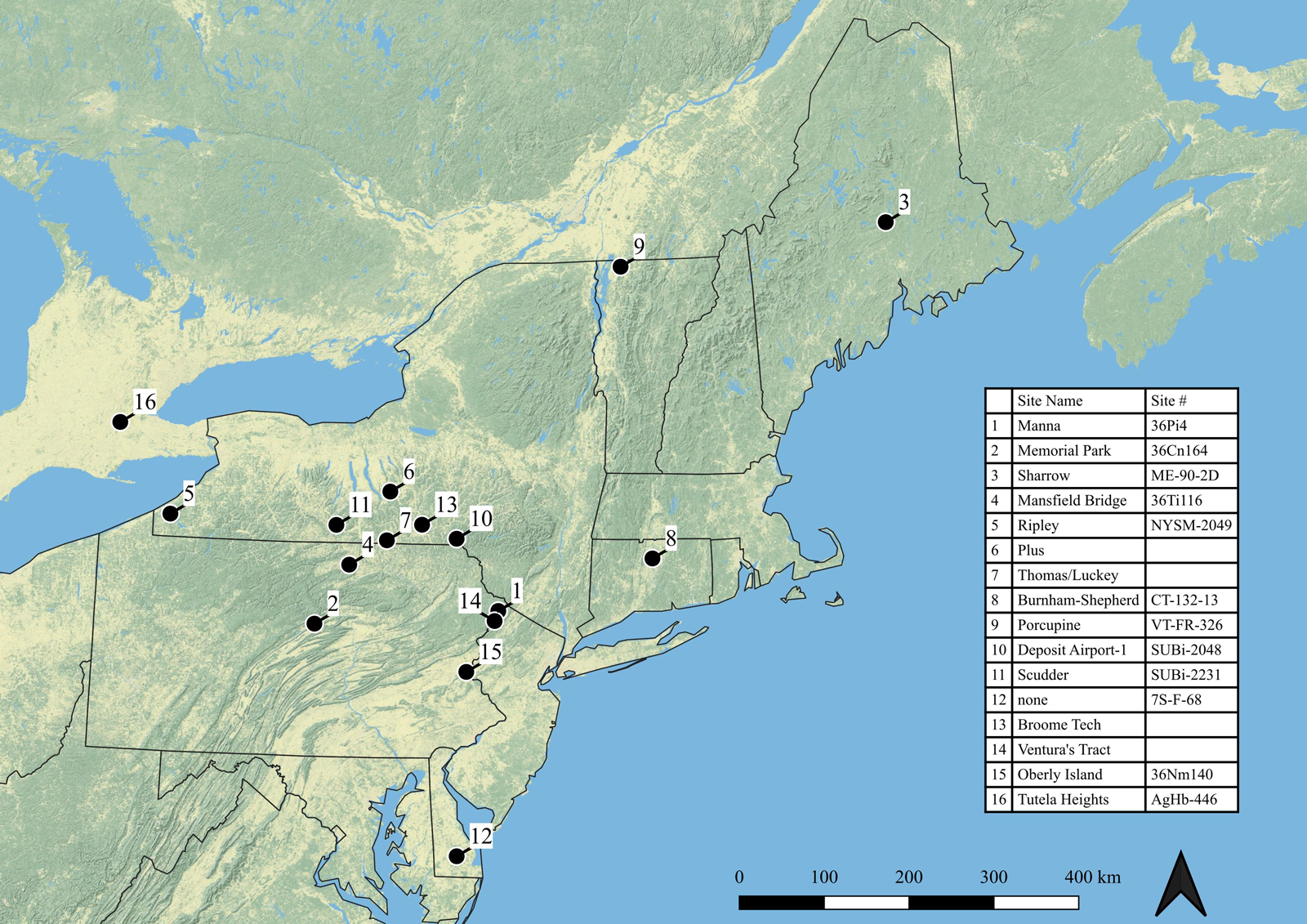
Figure 1. Archaeological sites from northeastern North America with EAC cultigens. Sites in the Monongahela River, Allegheny River, and Ohio River drainages in western Pennsylvania are not included.
Table 1. Archaeological Sites from Northeastern North America with Eastern Agricultural Complex Crops and Associated Dates.
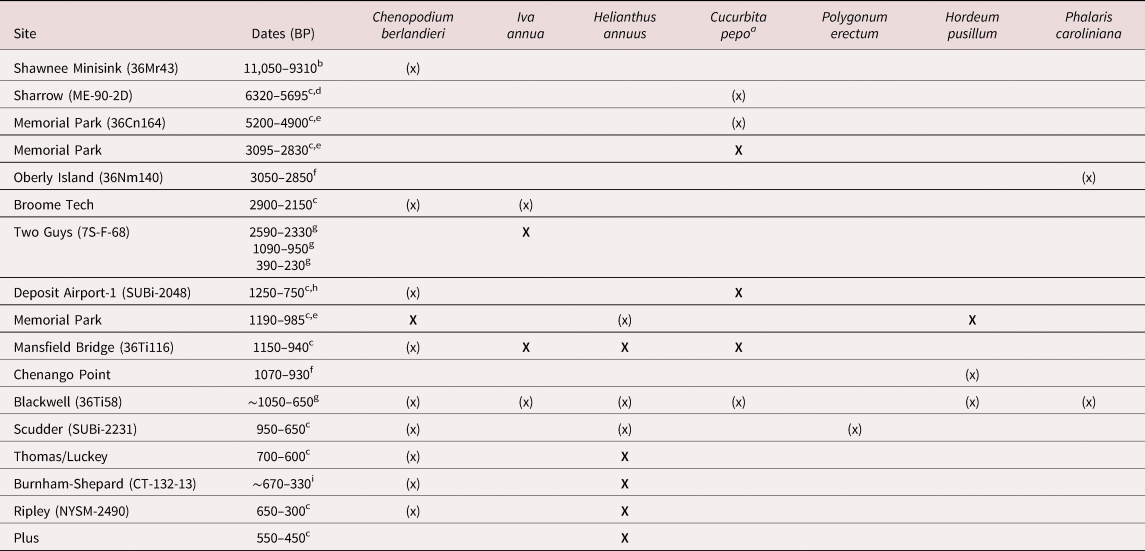
Note: X = domesticated; (x) = not domesticated
a Late Woodland sites with only Cucurbita pepo and no other EAC plants are excluded from this table, given that squash is commonly identified with maize from these sites (Hart Reference Hart and Hart2008; Hart and Asch Sidell Reference Hart and Sidell1997; Hart et al. Reference Hart, Thompson and Brumbach2003, Reference Hart, Brumbach and Lusteck2007; Thompson et al. Reference Thompson, Hart, Brumbach and Lusteck2004).
b Gingerich Reference Gingerich and Gingerich2013:Table 9.8.
c Asch Sidell Reference Asch Sidell and Hart2008:Table 3.3.
d Petersen and Asch Sidell Reference Petersen and Sidell1996:689.
e Hart and Asch Sidell Reference Hart and Sidell1997:528.
f Stewart Reference Stewart2018:Table 27.
g Messner Reference Messner2008:309–310; LeeDecker et al. Reference LeeDecker, Koldehoff and Holt1996.
h Asch Sidell Reference Asch Sidell, Knapp and Versaggi2002:120–121.
i Bendremer et al. Reference Bendremer, Kellogg and Largy1992 (no exact radiocarbon dates are provided, just an approximate age).
This article reports on the identification EAC cultigens from Middle and Late Woodland contexts at Manna (36Pi4), part of a large Lenape agricultural village located in the Upper Delaware Valley (UDV). The Manna assemblage provides evidence for cultivation of EAC plants by the Lenape as part of their broader agricultural systems. The presence of these plants at Manna complicates assumptions about Lenape plant cultivation systems and raises questions about the introduction of these plants into the region.
Eastern Agricultural Complex Plants in the Northeast
The Lenape and other Indigenous people in the Northeast consumed or used chenopodium since the Paleoindian period, with chenopodium identified in hearths at Shawnee Minisink (Gingerich Reference Gingerich2011, Reference Gingerich and Gingerich2013). Squash rinds found at Memorial Park along the Susquehanna River in Pennsylvania and at Sharrow in Maine were both dated to the Middle Archaic, although neither specimen is clearly domesticated (Table 1; Figure 1; Asch Sidell Reference Asch Sidell and Hart2008; Hart and Asch Sidell Reference Hart and Sidell1997:528; Petersen and Asch Sidell Reference Petersen and Sidell1996:689). Whether the range of these squashes had expanded naturally or through human intervention is still debated (Monaghan et al. Reference Monaghan, Lovis and Egan-Bruhy2006; Smith Reference Smith1989, Reference Smith2007). Domesticated squash was present in the region by the Late Archaic, with domesticated squash rind recovered from Memorial Park (Table 1; Hart and Asch Sidell Reference Hart and Sidell1997).
Beginning in the Early Woodland (ca. 900 BC–0 AD) and extending through the Late Woodland, chenopodium becomes more common in the archaeological record, although most of the populations examined do not appear to be from domesticated plants (Table 1; Asch Sidell Reference Asch Sidell and Hart2008; Belcher et al. Reference Belcher, Williams and Mueller2023). Throughout ENA, two types of domesticated chenopodium have traditionally been identified: red and pale morphs. Red morphs have smooth seed coats with testa thicknesses of less than 21 μm, truncated margins, and prominent beaks (Belcher et al. Reference Belcher, Williams and Mueller2023:557). Red morphs are hereafter referred to as thin testa phenotypes or morphs, given that color cannot be observed in carbonized specimens. Pale morphs completely lack the hard, black outer epiderm, with only the thinner, translucent inner epiderm remaining (Smith and Yarnell Reference Smith and Yarnell2009:6564). In contrast, wild chenopodium have black seed coats with testa thicknesses from 40 to 60 μm, are rounded to biconvex margins, and have alveolate patterning (Smith Reference Smith1985:59–60). The black, wild phenotypes are hereafter referred to as thick testa morphs or phenotypes. Previously, archaeologists had assumed that between 1% and 3% of the seeds on wild chenopodium plants would display the thin testa phenotype (Asch and Asch Reference Asch and Asch1977, Reference Asch, Asch and Ford1985; Smith Reference Smith1985). Consequently, the presence of a few domesticated, thin testa morphs has long thought to been indicative of Indigenous cultivation of domesticated chenopodium (Asch Sidell Reference Asch Sidell and Hart2008). More recently, surveys of modern, free-living stands of chenopodium have found that between 15% and 34% of seeds in these plants featured the thin testa morphs thought to be indicative of domesticated chenopodium (Belcher et al. Reference Belcher, Williams and Mueller2023; Halwas Reference Halwas2017). Common experimental garden research by Belcher and colleagues (Reference Belcher, Williams and Mueller2023) has further shown that after a single year of cultivation, 50% or more of the seeds produced by chenopodium plants featured thin testas. Furthermore, they found that the optimal conditions for the carbonization of thin testa morphs were different from those for the carbonization of thick testa morphs, although thin testa morphs on the whole were more likely to be fully carbonized and well preserved (Belcher et al. Reference Belcher, Williams and Mueller2023: 64).
The majority of the chenopodium populations identified in NENA could have been produced by wild stands of chenopodium (Asch Sidell Reference Asch Sidell and Hart2008:Table 3-3). However, two sites contained likely domesticated populations of chenopodium: Tutela Heights and Memorial Park (Crawford et al. Reference Crawford, Lytle, Williamson and Wojtowicz2019; Hart and Asch Sidell Reference Hart and Sidell1996).
The Tutela Heights assemblage, found in Ontario, originated from one pit feature dated to the Early Woodland (Figure 1). The majority of the chenopodium from the feature were thin testa morphs with relatively few thick testa morphs identified. Whether the specimens were grown locally or imported through trade into the Ohio River Valley could not be determined (Crawford et al. Reference Crawford, Lytle, Williamson and Wojtowicz2019). At Memorial Park, located along the Susquehanna River in Pennsylvania, pale morph chenopodium was identified along with little barley in contexts dating to the Late Woodland (Hart and Asch Sidell Reference Hart and Sidell1996). Other sites have been argued to have domesticated chenopodium based on the identification of few domesticated phenotypes within an assemblage (Asch Sidell Reference Asch Sidell, Knapp and Versaggi2002, Reference Asch Sidell and Hart2008). However, the proportion of domesticated-type chenopodium seeds identified in these assemblages is well within the range that wild plants produce, indicating that these do not represent domesticated populations (Belcher et al. Reference Belcher, Williams and Mueller2023; Halwas Reference Halwas2017).
Although only two truly domesticated populations of chenopodium have been identified in the paleoethnobotanical record of NENA, most undomesticated chenopodium seeds do not neatly fit into either domesticated or wild phenotypes. These intermediate-type seeds feature testa thicknesses above 21 μm but below 40 μm and rounded margins (Asch Sidell Reference Asch Sidell, Knapp and Versaggi2002, Reference Asch Sidell and Hart2008; Reference Asch Sidell and Michael Stewart2015; Crawford et al. Reference Crawford, Lytle, Williamson and Wojtowicz2019). Some have argued these intermediate seeds represent a cultivated but not yet domesticated chenopodium population (Fritz Reference Fritz and Gremillion1997; Gremillion Reference Gremillion1993; Halwas Reference Halwas2017). However, Belcher and colleagues (Reference Belcher, Williams and Mueller2023:564–565) argue for the retirement of the term “intermediate” because they contend that domesticated chenopodium should be measured based on a lack of seed polymorphism rather than testa thickness. Additionally, measurements of seed coat thickness and morphology indicate that there is no clear break between thin and thick phenotypes and that they instead occur on a gradient (Belcher et al. Reference Belcher, Williams and Mueller2023; Halwas Reference Halwas2017).
Along with chenopodium, marshelder begins to appear in the archaeological record during the Early Woodland, although only one site had domesticate-sized achenes. Marshelder is not considered native to NENA, with wild stands found in a single county each in Pennsylvania, Massachusetts, and Maine (Cullina et al. Reference Cullina, Connolly, Sorrie and Somers2011; Rhoads and Klein Reference Rhoads and Klein1993; Rhoads and Block Reference Rhoads and Block2007; Richards et al. Reference Richards, Hyland and Eastman1983). In these locations, marshelder is considered to be a waif, or a previously cultivated plant introduced by people that has become naturalized (Cullina et al. Reference Cullina, Connolly, Sorrie and Somers2011; Weiland and Gremillion Reference Weiland and Gremillion2018). Consequently, all marshelder found archaeologically in NENA is considered cultivated or introduced by Indigenous people (Asch Sidell Reference Asch Sidell and Hart2008). Squash is also identified more frequently after 1000 BC (Hart Reference Hart and Hart2008; Hart and Asch Sidell Reference Hart and Sidell1997; Hart et al. Reference Hart, Thompson and Brumbach2003, Reference Hart, Brumbach and Lusteck2007; Thompson et al. Reference Thompson, Hart, Brumbach and Lusteck2004).
Sunflower was part of Indigenous Algonquian maize-agricultural systems (e.g., Grant Reference Grant1907), and domesticated sunflower has been identified from five Late Woodland components in NENA (Asch Sidell Reference Asch Sidell and Hart2008:Table 3-3; Bendremer et al. Reference Bendremer, Kellogg and Largy1992). Sunflower has also been documented on Late Woodland sites in Ontario, with data suggesting that domesticated sunflower achene size is smaller than for other areas (Crawford and Smith Reference Crawford, Smith and Minnis2003). Domesticated and ruderal-sized marshelder have also been identified in Late Woodland components from three sites in the region (Table 1).
Erect knotweed, little barley, and maygrass are not commonly identified on archaeological sites in NENA. Despite being native to the region, erect knotweed has only been reported at one site represented by a single specimen (Asch Sidell Reference Asch Sidell and Hart2008:43). Aside from Memorial Park, little barley has only been identified via starch grain analysis on two sites (Messner Reference Messner2008:299). Maygrass was only identified at one Late Woodland site (Gardner Reference Gardner1992; Messner Reference Messner2008:309).
Evidence for the cultivation of EAC by the Indigenous people of NENA is incredibly scarce. Only three sites report more than two crops in their domesticated form or outside their native range (Table 1). My analysis of paleoethnobotanical remains at Manna indicates that the Lenape cultivated EAC plants throughout the Late Woodland as part of their maize-centered agricultural systems.
Manna (36Pi4)
Manna is a deeply stratified, multicomponent Lenape farmstead situated on the first terrace of the Delaware River, and it is a contributing site to the Minisink National Historic Landmark (NHLM)—an extensive Late Woodland Lenape village. Manna was excavated by a Temple University field school under the direction of R. Michael Stewart from 2003 for 2005 (Stewart Reference Stewart2015). A total of 46 1 × 1 m units were excavated across two seasons, with 72 features identified (Stewart Reference Stewart2015).
The earliest Lenape usage of the site predates 2880 ± 20 BP (UGAMS-59860; nutshell; δ13C = −22.96%), although paleoethnobotanical evidence indicates that the site was likely only occupied seasonally and focused on mast nut and fruit harvesting and processing until the end of the Middle Woodland (Reamer Reference Reamer2023). At the end of the Middle Woodland (1440 ± 20 BP), the Lenape began cultivating maize and were likely living in the Minisink NHLM year round (Reamer Reference Reamer2023; Stewart Reference Stewart2015:Table 17). During the Late Woodland, the Lenape used Manna primarily for farming maize and other crops, gathering mast nuts and fleshy fruits, and processing their harvests. Of the 72 features recovered, 60 dated to the Late Woodland. These included four hearths or firepits, one earth oven, four storage pits, 37 postmolds, and nine features of indeterminate function (Reamer Reference Reamer2023). No house patterns were discernible from the postmolds recovered, although temporary structures may have existed (Stewart Reference Stewart2015). The lithic and ceramic artifacts recovered from Late Woodland deposits also indicate a primary focus on plant processing activities (Stewart Reference Stewart2015). Based on the available evidence, the Lenape probably did not live at Manna year round and only used the area during the spring through fall months for agricultural production and harvesting (Reamer Reference Reamer2023). Manna also probably flooded regularly during the Late Woodland, which would preclude year-round habitation (Stinchcomb et al. Reference Stinchcomb, Messner, Driese, Nordt and Stewart2011).
Stewart (Reference Stewart2015) employed comprehensive flotation sampling to recover macrobotanical remains, allowing for detailed analysis of plant usage throughout the Lenape inhabitation. Samples were processed using a Siraf-type floatation machine. Soil samples of 2 L were taken from nonfeature contexts for each arbitrary 5 cm excavation level. Feature matrixes were floated in their entirety, but sample volume was not recorded, so overall flotation volume cannot be ascertained (Stewart Reference Stewart2015:86). I standardized counts using charcoal weight. Seventeen samples were previously analyzed by Nancy Asch Sidell (Reference Asch Sidell and Michael Stewart2015) from Feature 89, a hearth dated to 840 ± 70 BP (Table 2; Stewart Reference Stewart2015:118, Table 17). Following the methodology outlined by Pearsall (Reference Pearsall2015), I analyzed an additional 390 flotation samples in the North American Archaeology Laboratory at the University of Pennsylvania, for a total of 407 samples. A total of 2,250 maize fragments, 5,133 nutshell fragments, and 2,069 seeds and squash rinds were identified. Of the 2,069 seeds and squash rinds, 543 were from EAC species (Table 2).
Table 2. Flotations Contexts from Manna with Eastern Agricultural Complex Cultigens.
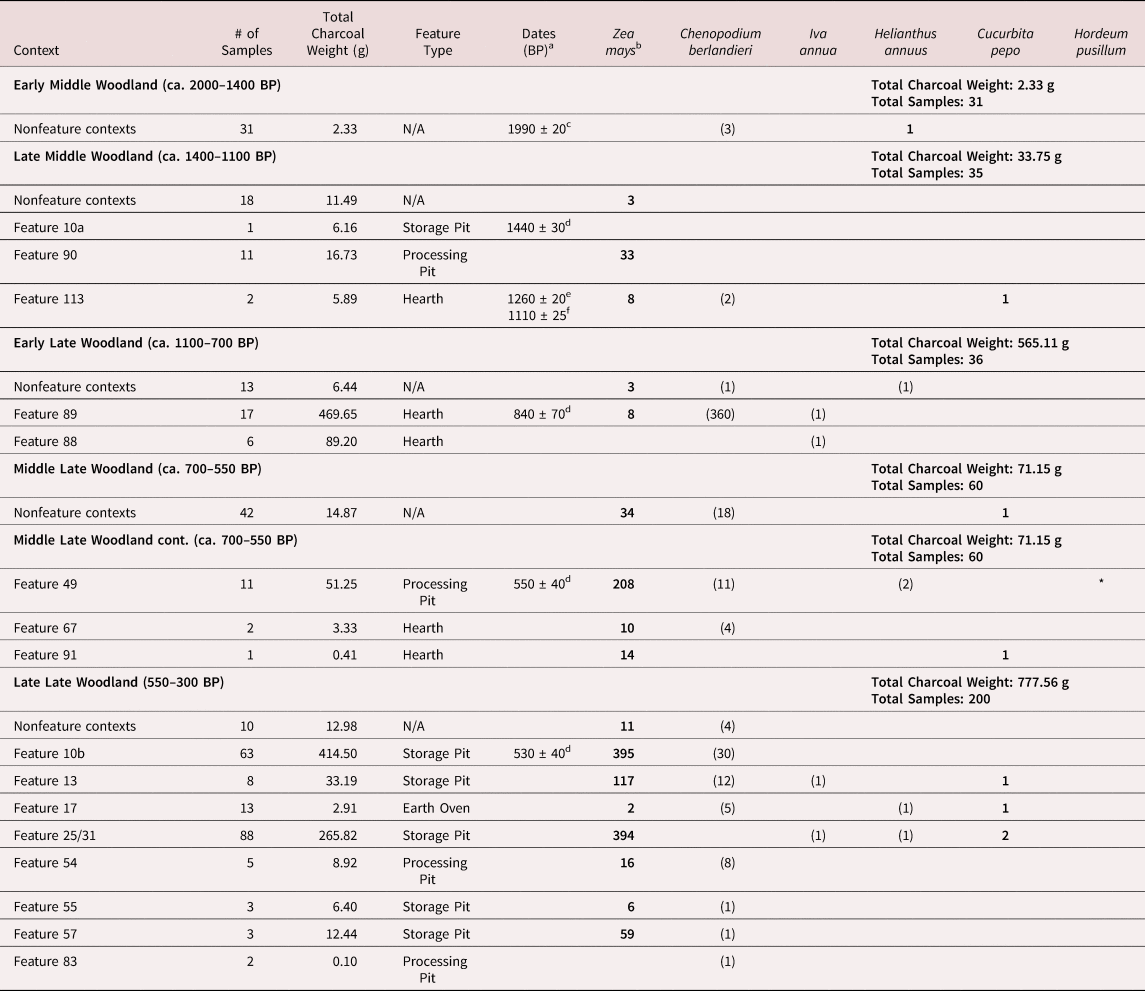
Notes: Only samples with EAC plants are listed. Numerals in bold = domesticated; numerals in parentheses = not domesticated; * = starch grain only.
a Dates only listed for features that have been AMS assayed.
b Possible maize kernels and cupules were not included in these counts.
c UGAMS-59855, nutshell, δ13C = −24.58%.
d Stewart Reference Stewart2015:Table 17.
e UGAMS-59856, nutshell, δ13C = −25.53%.
f UGAMS-133843, unknown, δ13C = −27.91%.
Chenopodium / Lamb's Quarters (Chenopodium berlandieri, n = 461)
Chenopodium was the most abundant EAC seed identified at Manna. The majority of the chenopodium seeds (n = 360) were identified by Asch Sidell (Reference Asch Sidell and Michael Stewart2015) and could not be assessed for domestication, given that most were destroyed when rehoused into plastic bags by the National Park Service from the original vials (Asch Sidell, personal communication 2022). Luckily, two vials were still intact and could be examined for signs of domestication. A total of 15 chenopodium seeds from Feature 89 were examined to document testa thickness and margin shape (Table 3). Six seeds were examined by Asch Sidell (Reference Asch Sidell and Michael Stewart2015:375, Table 5), and nine were examined by the author under a Keyence microscope in the Penn Paleoecology Lab at 200–300× magnification. Given that only specimens retaining part of their testa were examined, other samples with identified chenopodium did contain a sufficient number of specimens to adequately assess the population.
Table 3. Chenopodium Measurements from Feature 89 at Manna.

a Measured by Asch Sidell Reference Asch Sidell and Michael Stewart2015:Table 5.
Similar to other Late Woodland chenopodium populations in NENA (Asch Sidell Reference Asch Sidell and Hart2008:40–42), most chenopodium from Manna featured rounded-to-biconvex margins and intermediate testa thicknesses (average = 28.1 μm; Figure 2a–b; Table 3). Three specimens were more similar to the traditionally defined domesticated black morphs with truncated margins and testa thicknesses below 21 μm (Figure 2c–e; Table 3: Specimens 10, 14, 15). The seeds analyzed from Manna do not indicate a domesticated population. Only 30% of the examined seeds featured thin testas, which is within the range of wild plants and populations identified in broader ENA and southern Manitoba (Belcher et al. Reference Belcher, Williams and Mueller2023; Halwas Reference Halwas2017). However, the overall low average testa thickness could indicate a cultivated population of chenopodium. Similar populations between domesticated and wild populations observed elsewhere in ENA and southern Manitoba are speculated to have been from cultivated but not domesticated populations (Fritz Reference Fritz and Gremillion1997; Gremillion Reference Gremillion1993; Halwas Reference Halwas2017). Further research on chenopodium populations throughout NENA is needed to determine if the Indigenous people of the region were cultivating chenopodium, and therefore unintentionally selecting for thinner testas to improve germination rates, or if wild chenopodium plants in the region have thinner testas as observed elsewhere (Halwas Reference Halwas2017). No chenopodium seeds from deposits postdating Feature 89 could be examined for testa thickness to determine how chenopodium testa thickness changed throughout the Late Woodland. The Lenape appear to have harvested less chenopodium, whether wild or cultivated, throughout the Late Woodland, but chenopodium was still identified in features across all Late Woodland components, indicating that the Lenape were still utilizing the plant even as maize became a more important food source (Table 3).

Figure 2. Chenopodium samples from Manna: (a) Specimen 11 top view showing reticulated testa pattern; (b) Specimen 11 side view showing seed coat thickness and round to biconvex margin; (c) Specimen 15 top view showing smoother testa pattern; (d) Specimen 15 side view showing truncated margins; (e) Specimen 15 view showing seed coat thickness measurement locations. (Color online)
Sunflower (Helianthus annuus, n = 6)
Six sunflower kernels and achenes were identified at Manna. One kernel from a nonfeature context dated to 1990 ± 20 BP (UGAMS-59855; nutshell; δ13C = −24.58%) was clearly domesticated. The kernel measured 4.824 mm long, and it was 1.921 mm wide at its widest extant point, with a size index (L × W) of 9.267 (Figure 3). Carbonized wild kernels measured by Smith (Reference Smith2014:Figure 9) had maximum lengths of 4.2 mm and size indexes below 8. The kernel from Manna is most similar to domesticated, carbonized kernels from Late Archaic assemblages in Illinois and Tennessee (Smith Reference Smith2014), although Crawford and Smith (Reference Crawford, Smith and Minnis2003) note that domesticated sunflower in NENA and Ontario were smaller than similarly dated specimens. Given that the domesticated kernel was from a nonfeature context and not directly dated, the early date should be regarded with some skepticism. However, because no domesticated sunflower has previously been reported in the UDV, the finding of any domesticated sunflower is still significant (Stewart Reference Stewart2018). The other sunflower kernels from Manna were too fragmentary to obtain accurate measurements. Sunflower kernels and achenes were present in components from throughout the Late Woodland, indicating that the plant was likely part of Lenape maize agricultural systems, as has been observed throughout NENA (Asch Sidell Reference Asch Sidell and Hart2008; Bendremer et al. Reference Bendremer, Kellogg and Largy1992; Crawford and Smith Reference Crawford, Smith and Minnis2003).

Figure 3. Early Middle Woodland domesticated sunflower kernel. (Color online)
Squash/Gourd Rind Fragments, n = 7
Squash was only identified at Manna from rind fragments. Lack of squash seeds in the botanical assemblage is not uncommon because the high oil and moisture content of the seeds inhibit carbonization, and squash seeds were frequently consumed (King Reference King and Ford1985; Simon Reference Simon2011; Smith Reference Smith2007). The earliest rind fragments from Manna came from Feature 113, which dated to 1260 ± 20 BP (UGAMS-59856; nutshell; δ13C = −25.53). The late Middle Woodland rind was identifiable as a cucurbit based on the presence of epidermal cystoliths. Viewed in cross section, the inner rind cells were orderly and had an isodiametric shape, indicating that the fragment is Cucurbita pepo (Asch and Asch Reference Asch, Asch and Ford1985). The other squash rinds from Manna were not examined in cross section but are assumed to also be Cucurbita given that no bottle gourds have been documented in the UDV (Stewart Reference Stewart2018). Ethnobotanical and ethnohistoric accounts also do not document any Lenape cultivation of bottle gourds. The rind from Feature 113 is the earliest documented squash in the UDV. Lenape cultivation of squash is not surprising given that squash cultivation was widespread throughout NENA and was previously documented in the UDV (Stewart Reference Stewart2018:Table 27).
Marshelder (n = 4)
Four marshelder kernels were recovered from Late Woodland contexts at Manna. The Feature 89 kernel measured 2.2 × 1.8 mm with an estimated achene size of 3.16 × 2.55 mm, both corrected for carbonization effects (Asch Sidell Reference Asch Sidell and Michael Stewart2015:374). Other kernels were of similar size, but unfortunately, they were fragmented before detailed measurements could be obtained (Messner Reference Messner2011). Modern, wild marshelder achenes have an average size of 2.8 × 2.2 mm and range from 1.6 to 4.6 mm in length, whereas domesticated populations from Illinois were between 3.0 and 8.6 mm in length (Asch and Asch Reference Asch, Asch and Ford1978; Asch Sidell Reference Asch Sidell and Michael Stewart2015:374). The baseline for domesticated marshelder achene length is 4.0 mm (Smith Reference Smith1989). Although the samples from Manna were not domesticated, marshelder is not native to the UDV, which indicates that the kernels were likely cultivated by the Lenape or traded into the region. Marshelder was identified in both early Late Woodland and late Late Woodland components, indicating possible cultivation throughout the Late Woodland. Alternatively, previously cultivated marshelder could have continued growing without Lenape interference, given its current status as a waif in parts of NENA.
Erect Knotweed and Little Barley
Although various species from the Polygonaceae family were identified at Manna, none were conclusively identified as erect knotweed. Possible little barley starch was recovered from a tabular grinding stone found in Feature 49 (Messner Reference Messner2008:85, 2011). However, the starch grains also could have originated from wild rye (Elymus sp.). No little barley grains were recovered.
Discussion and Conclusion
The Manna paleoethnobotanical assemblage indicates that the Lenape were consuming if not cultivating at least four EAC cultigens alongside maize and beans throughout the Late Woodland. Evidence for possible chenopodium cultivation includes the generally thinner testas observed in Feature 89, although this could be unique to the population the Lenape were exploiting. Regardless of whether the Lenape were cultivating chenopodium, they were consuming the seeds from at least the Middle Woodland through the end of the Late Woodland. Marshelder is not native to the UDV and was recovered from early Late Woodland and late Late Woodland components, indicating possible continued cultivation throughout the Late Woodland. Sunflower was previously assumed to have been part of Lenape agricultural systems, but domesticated sunflower had not previously been recovered in the region. Although squash had previously been recovered in the UDV from Late Woodland contexts, the paleoethnobotanical assemblage from Manna shows that the Lenape were cultivating squash by the Middle Woodland (Table 2; Stewart Reference Stewart2018:Table 27). The presence of EAC cultigens from throughout the Late Woodland complicates previous assumptions about Lenape and other Indigenous agricultural systems in NENA.
Models of Indigenous agricultural systems in NENA during the Late Woodland generally focus only on the cultivation of maize, beans, squash, and occasionally sunflower (Crawford and Smith Reference Crawford, Smith and Minnis2003; Hart Reference Hart and Hart2008; Hart and Lovis Reference Hart and Lovis2013). All other plants, including chenopodium and marshelder, are generally considered to be wild resources (Hart and Lovis Reference Hart and Lovis2013). However, my analysis of the paleoethnobotanical remains from Manna indicate that Lenape agricultural systems were more diverse than previously acknowledged. Lenape agricultural systems were probably more similar to those described across ENA after circa AD 1300, with maize as the staple crop supplemented by EAC cultigens (Fritz Reference Fritz2019; Hart and Lovis Reference Hart and Lovis2013; Mueller et al. Reference Mueller, Fritz, Patton, Carmody and Horton2017). The Lenape may have grown sunflower, chenopodium, and marshelder along the edges of their maize fields during the early Late Woodland, as described by Fritz (Reference Fritz2019) at Cahokia around the same time. The possibility of similar cultivation practices during this time period could indicate exchange of agricultural knowledge across ENA even without the movement of other cultural practices.
Even as the Lenape increased their cultivation of maize throughout the Late Woodland, they still consumed and possibly cultivated sunflower, marshelder, and chenopodium. The Lenape's probable cultivation of EAC plants during the late Middle and early Late Woodland emphasizes the need for more systematic paleoethnobotanical research in NENA to better understand Indigenous agricultural systems and move beyond maize-centric assumptions.
Future research is also needed to determine when the Lenape, and other Indigenous groups in NENA, first began cultivating both EAC plants and maize. The earliest possible EAC cultigen at Manna was the domesticated sunflower kernel recovered from early Middle Woodland contexts. I could not directly date this specimen due to the terms of my loan with the NPS, and I therefore cannot safely conclude whether the kernel is from the early Middle Woodland or intrusive from a later deposit. Maize may have been introduced into the region as early as 2200 BP, although further direct dates are needed to confirm this as well (Hart et al. Reference Hart, Brumbach and Lusteck2007; Stewart Reference Stewart2018). Future paleoethnobotanical analysis will help to elucidate whether the Lenape and other NENA Indigenous groups adopted maize and EAC cultigens as a package of cultivated plants, whether the plants were introduced separately, or whether cultivation of EAC plants emerged independently in NENA. Regardless, the cultivation of maize with EAC plants shows that Lenape agricultural systems were more similar to those observed in broader ENA than previously realized.
Acknowledgments
I thank the Delaware Tribe of Oklahoma, the Delaware Nation of Oklahoma, the Stockbridge-Munsee Community, and the National Park Service DEWA offices for allowing access to the Manna collections. I thank Kathleen Morrison and Moriah McKenna for allowing me to use the Penn Paleoecology Laboratory. Thank you to Anna Fuller Graham and Nancy Asch Sidell for their help in identifying plant specimens from the Manna assemblage. Thanks to Megan Kassabaum, Alexandria Mitchem, and the anonymous reviewers for providing comments to improve the article.
Funding Statement
This research was carried out with funding from the Louis J. Kolb Society at the Penn Museum.
Data Availability Statement
All data is on file with the National Park Service DEWA Office in Bushkill, Pennsylvania, or available by contacting the author. The Manna dataset discussed here is permanently curated at the same facility.
Competing Interests
The author declares none.







 Open access
Open access