


Introduction
The introduction of exotic species is largely seen as deleterious for ecosystems and for the survival of native species (Williamson, Reference Williamson1996). However, in spite of the general concerns about introductions of exotic species there are contrary arguments (in the scientific and popular press) that have developed the idea that introduced species could increase local biodiversity and provide some ecosystem services (Sax & Gaines, Reference Sax and Gaines2003; Warren et al., Reference Warren, Fleishman, Mcgeoach, Mcdonall, Chown and MacNally2004; Schutzenhofer & Valone, Reference Schutzenhofer and Valone2006). The argument is theoretically strong and could influence management decisions regarding recently settled, potentially invasive, alien species.
The large amount of data available on the impacts of alien invasive species shows that decision makers should avoid waiting for the occurrence of local impacts before deciding to eradicate species (Lobos & Jaksic, Reference Lobos and Jaksic2005). However, interactions between introduced and native species belonging to the same family or the same functional group have been little studied, particularly in vertebrates. The few studies have generally underlined the negative impacts of introduced vertebrates (Mountainspring & Scott, Reference Mountainspring and Scott1985; Bennet, Reference Bennet1990; Gurnell et al., Reference Gurnell, Wauters, Lurz and Tosi2004; Schumacher & Parrish, Reference Schumacher and Parrish2005; Baxter et al., Reference Baxter, Fausch, Murakami and Chapman2007) but some showed no real impacts (Moffat & Minot, Reference Moffat and Minot1994; Rissler et al., Reference Rissler, Barber and Wilbur2000; Kawakami & Higuchi, Reference Kawakami and Higuchi2003). In the light of the latter results it has been postulated that the decision to control ‘new but nice’ introduced species could be delayed. For example, the proposed eradication of the alien sacred ibis Threskiornis aethiopicus was postponed in western France following the suggestion that the species could have a positive impact on a closely related species, the Eurasian spoonbill Platalea leucorodia. The argument is that the recently settled colonies of ibises could attract and stabilize the unstable colonies of spoonbills (Marion, Reference Marion2006). To our knowledge this is the first time that such a direct positive impact between an introduced and a native vertebrate has been proposed. This suggestion, which has had an important influence on decision making over the ibis in France, needs to be demonstrated or invalidated.
We analysed the spatial relationships between the native spoonbill and the introduced sacred ibis during the breeding season in a large swamp where the two species can nest without spatial constraints, the Grand-Lieu Lake in western France. These wading birds are large, white species of the Family Threskiornithidae and have some similarities in feeding and nesting behaviour. The spoonbill breeds in Eurasia and Africa (Triplet et al., Reference Triplet, Overdijk, Smart, Nagy, Schneider-Jacoby and Karauz2008) and is not categorized as globally threatened (it is categorized as Least Concern on the IUCN Red List; BirdLife International, 2009) but is considered Vulnerable in France (Dubois et al., Reference Dubois, Le Maréchal, Olioso and Yésou2008). This species began to nest at Grand-Lieu Lake in 1981 and the number of pairs increased to 77 in 2006. The sacred ibis has a sub-Saharan African origin and does not occur naturally in Europe. Birds escaped from a zoo and began to breed on Grand-Lieu Lake in 1994 (up to 191 pairs in 2005). Since 2001 this species has been of concern for the managers of protected areas in France because of several cases of it predating on broods of species of conservation concern (Yésou & Clergeau, Reference Yésou and Clergeau2005; Clergeau & Yésou, Reference Clergeau and Yésou2006). Colonies of these two gregarious species also occur, singly or together, in several other marshes and islands in the same region.
To analyse the relationships between the two species and any attraction between them, we focused on their possible interactions during colony settlement. We analysed both the temporal and spatial patterns of nest distribution and the choice of nest neighbourhood by settling spoonbills.
Methods
The study was conducted on the c. 6,300 ha Grand-Lieu Lake, western France, which includes a nature reserve of c. 2,700 ha. From aerial photographs and upon advice from reserve staff we first identified the potential nesting areas for both species, corresponding to the extent of the habitats where colonies have been observed. This area (Fig. 1), covering 1,365 ha, is a relatively homogeneous swamp with reed Phragmites australis, willows (Salix atrocinerea and Salix fragilis) and alder Alnus glutinosa. Other colonial wading birds breed in this area, especially the grey heron Ardea cinerea, but their nests occur throughout the swamp.
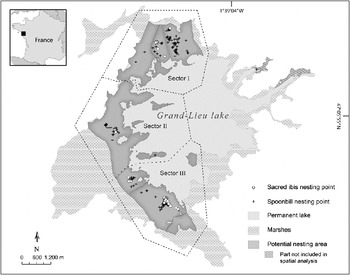
Fig. 1 Locations of nests of the native spoonbill Platalea leucorodia and the introduced sacred ibis Threskiornis aethiopicus during 2002–2006 in the study area on the western side of Grand-Lieu Lake. The area was divided into three sectors for analysis. The inset indicates the location of the main map in western France.
We used the census data collected by the reserve staff during 2002–2006. Censuses were conducted every 5 days from 1 April to 30 June and covered the whole area studied. The locations of nests, which all occurred in trees, were recorded with a global positioning system and imported into the geographical information system ArcInfo v. 9.2 (ESRI, Redlands, USA). Ibises can breed very close to each other (< 1 m between nest centres) and therefore clusters of up to 17 nests were given the same coordinates. For each nest the date at which the first egg was laid was either observed or, more often, calculated from the observed age of the pulli (S. Reeber, unpubl. data). Beginning of egg laying is a good proxy for the date when each breeding pair settles in the colony because, in both species, the breeders settle and begin to build their nest only 6–12 days before laying (S. Reeber & P. Yésou, unpubl. data). This method ensures that each of the nests included in the analyses was occupied. We examined the breeding chronology of both species separately for each of the 5 study years, using the cumulative number of nests per 5-day period (i.e. the time between consecutive surveys).
The spatial distribution of nests was examined using ArcGis. The relationships between ibises and spoonbills were analysed with a grid analysis (Fortin & Dale, Reference Fortin and Dale2005). The grid contained 2,674 square cells of width 80 m; this width was based on the highest annual median value (39.75 m) of nearest neighbour distances between ibises and spoonbills. The history of occupied cells was examined for each year. We noted cells used by a single species or by both species, and whether the occupied cell was occupied in the previous years and by which species. We thereafter examined the spatial difference between the distributions of colonies in three sectors of relatively equal area (Fig. 1). We used ArcGis to calculate mean centres, in geographical coordinates, of all nesting points for each species in each of the three sectors per year. As a nesting point can represent more than one nest for the ibis (1–17, mean = 3.6), we weighted each nesting point by the number of nests there. To compare the dispersion of nesting points around their mean centre we calculated the standard distance, which is the standard deviation of the distance of each point from the mean centre.
To examine any potential influence of nesting ibises on spoonbill settlement we analysed the choice of neighbourhood for each new spoonbill nest (Dixon, Reference Dixon1994). We restricted our analysis to sectors and dates when ibises and spoonbills were already settled. For each spoonbill nest we identified both the nearest neighbouring nest (species and distance) and all the nests already occupied in neighbourhoods of radii of 50, 100 and 200 m.
Results
The number of spoonbill nests with eggs (mean 42.6 ± SD 9.7 per year) increased in 2005–2006 whereas the annual number of ibis nests with eggs (mean 142.8 ± SD 16.9 per year) was more variable. In all 6 years the period of nest initiation was longer in spoonbills (c. 95 days, mid February to end of May) than in ibises (c. 60 days, mid March to mid May). The chronology of the numbers of nests (Fig. 2) shows that spoonbills began to breed significantly earlier (by 15–25 days) than ibises in all 5 years (127 < df < 201, all t > 3.04, all P < 0.001). When the first ibis laid its first egg, between 31% (in 2005) and 72% (in 2003) of the spoonbills had already begun laying, a large proportion were advanced in incubation and a few already had young in the nest.

Fig. 2 Cumulative percentage of the number of spoonbill and ibis nests at Grand-Lieu Lake (Fig. 1) during 2002–2006. Censuses were every 5 days. The horizontal hatched line indicates the percentage of spoonbill nests that have their first egg when the first ibis nest has its first egg.
The map of nest locations shows the aggregated distribution of the colonies, mostly concentrated in the four parts of the swamp (Fig. 1) furthest from disturbance by people. Sector II was used for the first time by spoonbills in 2002 (16 nests), and was thereafter used every year, but ibis nests were not observed in this Sector prior to 2005. Of the 2,674 cells 113 (4.23%) were used by one of the two species at least once during the study and only 33 cells (1.23%) were used for more than 1 year. Although the number of ibis pairs was twice that of spoonbill pairs, the number of cells with ibis nests was significantly lower than those with spoonbills because ibises bred in more compact colonies than spoonbills (Table 1, and see below). Both species used, firstly, cells from which the other species was absent and, secondly, cells occupied by their own species. There was little site fidelity between years; both species nested mainly on trees not occupied in previous years (Table 1; χ2 tests, all P < 0.0002). In both analyses (1 year vs the previous year or 1 year vs all previous years), spoonbills settled more often in a cell previously used by conspecifics than ibises did.
Table 1 Number of square cells (of width 80 m, see text for details), of a total of 2,674 cells at Grand-Lieu Lake (Fig. 1), occupied by spoonbill Platalea leucorodia or ibis Threskiornis aethiopicus or both species in 1 year and occupied by spoonbill, ibis, both species or neither species in the previous year or in the previous 1–4 years.

The mean centre of the nests of each species in each sector varied between years, and distances between mean centres of the two species was 150–710 m. The standard distance deviations of ibis nests were 34–311 m and those of spoonbill nests were 63–771 m. The overall compactness of the distribution of ibis nests was twice that of spoonbills (Table 2).
Table 2 Standard deviations of distance (m) between nest locations and the mean geographical centre of spoonbill and ibis colonies, by sector, for 2002–2006, and F-tests of differences between species by sector and year. Empty cells indicate that the species was absent from that sector in that year.

*P < 0.05; **P < 0.001
The study of the distance to the nearest neighbour at the time of spoonbill settlement showed that the 120 spoonbill nests analysed from 2002 to 2006 settled significantly more often closer to a spoonbill nest (90.3%, varying from 77.8% in 2005 to 100% in 2003) than to an ibis nest. The mean distance between a new spoonbill nest and its nearest neighbour did not vary significantly, whether the neighbour was a spoonbill nest (33.13 m ± SD 30.69, n = 108) or an ibis nest (27.54 m ± SD 27.40, n = 12). The study of neighbourhoods of radii of 50, 100 and 200 m (Table 3) thus corroborates the spoonbill’s tendency to settle closer to conspecifics even if ibis nests are more abundant in the studied area.
Table 3 Percentage of spoonbills that nested in the neighbourhood (of radii of 50, 100 and 200 m) of ibis nests only, of ibis and spoonbill nests or of spoonbill nests only.

Discussion
The temporal and spatial patterns of nest distribution observed at Grand-Lieu Lake do not support the assertion that spoonbills are attracted to nest near ibises and consequently do not support the proposed role for the sacred ibis in the conservation of Eurasian spoonbills through facilitation of colony settlement. Analyses of spoonbill nests showed that in all years this species began to breed considerably earlier than the ibis, thus eliminating the possibility of any strong attraction of spoonbills towards ibises. Every year ibises began to nest > 15 days after the spoonbills. The ibises, which settle in a breeding site only a few days before they engage in nest building (authors, pers. obs. at various sites), were not yet present when a significant proportion of spoonbills had already begun to lay. Spatial analysis showed that the two species tended to concentrate in particular parts of the swamp, sometimes in small colonies of a few nests. However, the ibises were more gregarious than spoonbills and both species mostly nested in different trees each year.
Analyses, based on both grid and distance methods, corroborated the lack of interaction in nest location between the two species. The fact that the birds were more concentrated in some parts of the marshes, and particularly in three sectors, is best explained by their timid character. Both species avoid the border of the swamp, although the tree habitat there is similar to elsewhere in the swamp and suitable for nest building. The aggregations of nest sites in four areas, which gives the impression that ibises settle to breed in areas where spoonbills are also nesting, are in the most undisturbed parts of the swamp and the intervening areas (with no nesting) are close to the canals used by people. A similar preference for nesting in isolated locations is observed at other regional breeding sites (Herring & Gawlik, Reference Herring and Gawlik2008), corroborating the suggestion that the two species prefer secluded areas less subject to human disturbance. The more precise analyses of nest neighbourhood choice by spoonbills underlined the lack of any attraction towards ibises: 90% of spoonbills nested close to another spoonbill nest and, even considering a large radius and the higher frequency of ibis nests, newly arriving spoonbills preferentially chose the neighbourhood of other spoonbills.
Although our results do not support either a positive or negative effect of ibises on spoonbill nest timing or distribution, the assertion of a positive relationship between the two species had an important impact on decision making for the management of sacred ibises in France. It was one of the main reasons for postponing the proposed eradication project in 2005, which finally began only in 2008 after numerous discussions and several interventions by nature reserve managers. However, if the false assertion of a positive effect of ibises on spoonbill nesting had so easily influenced decision makers, it was also because the various bodies and people involved (e.g. general public, ornithologists and politicians) were not ready to accept control strategies for a species with an attractive appearance. The categorization of the sacred ibis as an invasive, exotic or alien species appeared not to be a sufficient point, indicating the need for better public information on the potential dangers posed by exotic animals and plants.
The impacts of the sacred ibis are known: in France it predates tern and heron broods (Clergeau & Yésou, Reference Clergeau and Yésou2006; Clergeau et al., Reference Clergeau, Reeber, Bastian and Yésou2010) and in South Africa cormorant eggs (Williams & Ward, Reference Williams and Ward2006), and there are also public health concerns (Epstein et al., Reference Epstein, Mckee, Shaw, Hicks, Micalizzi and Daszak2007) and urban nuisance (Martin et al., Reference Martin, French and Major2007) caused by Threskiornis molucca. Consequently, it would be reasonable to apply the precautionary principle, eradicating the first ibises that nest in any new region outside their African range. In 2008 the Andalusian authorities began an eradication process as soon as some ibises were observed in Doñana National Park (Carlos Gutiérrez Expósito, pers. comm.). One of the new challenges for conservation ecology is to avoid the settlement and establishment of newly introduced exotic species, in particular when these species have previously demonstrated negative impacts elsewhere. The recent examples of the zebra mussel Dreissena polymorpha in the USA (Zhu et al., Reference Zhu, Fitzgerald, Mayer, Rudstam and Mills2006), plecos fishes Hypostomus plecostomus in Mexico (Mendoza et al., Reference Mendoza, Contreras, Ramírez, Koleff, Alvarez and Aguilar2007) and the tamia squirrel Tamias sibiricus in France (Pisanu et al., Reference Pisanu, Marmet, Beaucournu and Chapuis2008), among others, illustrate cases of serious new problems that were not expected. In addition, the ‘tens rule’ (10% of introduced species settle and 10% of settled species became pests; Williamson, Reference Williamson1996), although it may be applicable to alien plants, may not apply to birds and mammals, for which the proportion of introduction events leading to problems seems to be 15–50% (Jeschke, Reference Jeschke2008; DAISIE, 2009). As prevention remains the most important tool, the role of zoos and the wildlife trade in the introduction of exotic animals such as the sacred ibis must not be underestimated (Clergeau & Yésou, Reference Clergeau and Yésou2006; Smith et al., Reference Smith, Behrens, Schloegel, Marano, Burgiel and Daszak2009).
Acknowledgements
We thank all the staff of the Grand-Lieu reserve for their contribution and C. J. Feare for his comments on the first draft of this article.
Biographical sketches
Philippe Clergeau’s research focuses on mechanisms for biodiversity conservation under anthropogenic pressures, and especially on the dynamics and impacts of alien vertebrates and on the development of urban biodiversity. He is the leader of multidisciplinary programmes on the role of ecological corridors. Damien Fourcy specializes in cartography and spatial analysis. Sébastien Reeber is a staff member of the society that manages the large nature reserve at Grand-Lieu Lake, and has a particular responsibility for bird censuses. Pierre Yésou is an ornithologist in a department responsible for management of hunting and wildlife in western France. He is a specialist in the birds of marine and freshwater environments.





