


Colorectal cancer (CRC) is one of the most frequently diagnosed malignancies worldwide, accounting for 10 % of all cancers and for approximately 20 % of all cancer-related deaths in developed countries( Reference Jemal, Center and DeSantis 1 ). Although there is a genetic component in CRC development, diet and other lifestyle factors are estimated to explain as much as 30–50 % of the global incidence of the disease( 2 ). According to the recent report from the World Cancer Research Fund and American Institute for Cancer Research (WCRF/AICR)( 2 , 3 ), there is convincing evidence that intake of red and processed meat increases the risk of CRC, whereas intake of dietary fibre is protective( Reference Murphy, Norat and Ferrari 4 ).
A variety of mechanisms have been proposed to link red and processed meat consumption and the risk of CRC( Reference Bastide, Pierre and Corpet 5 ). For red meat, in particular, it has been suggested that its high content of haem Fe is a substantial contributor( Reference Rijnierse A and de Wit 6 ). Red meat undergoes fermentation in the colon that might alter the microbiota composition and result in the production of potentially genotoxic products that could play a role in oncogenesis. These agents include N-nitroso compounds (NOC), a complex mixture of nitrite-derived products formed either in processed meat itself or endogenously in the human gut via bacterial metabolism. NOC are alkylating agents that generate DNA adducts in human colonocytes after high red meat consumption( Reference Lewin, Bailey and Bandaletova 7 ). We have recently shown that the level of the pro-mutagenic adduct O6-methyl-2-deoxyguanosine (O6MeG) is increased in murine colonocytes after consuming a diet high in red meat( Reference Winter, Nyskohus and Young 8 ). O6MeG is a known toxic and mutagenic base modification that, if unrepaired, can induce GC → AT transition mutations (typically found in the K-ras gene in human CRC)( Reference Lees, Harrison and Hall 9 ) and also recombination events or mutations in the form of sister chromatid exchanges( Reference Margison, Santibanez Koref and Povey 10 ). More recently, it has been suggested that high red meat consumption can increase the expression of certain oncogenic microRNA( Reference Humphreys, Conlon and Young 11 ).
Dietary fibre is a heterogeneous group of compounds, principally indigestible carbohydrates of plant origin that include NSP, starches that escape digestion in the small intestine (resistant starches, RS) and oligosaccharides. One possible mechanism for the reduction in the risk of CRC by dietary fibre is the production of SCFA via fermentation by the large-bowel microbiota( Reference Topping and Clifton 12 ). Of the major SCFA, butyrate is of particular interest as it appears to be the preferred metabolic substrate for colonocytes, and butyrate also promotes a normal cellular phenotype. In vitro studies with CRC cell lines have shown that butyrate induces apoptosis( Reference Medina, Edmonds and Young 13 ), reduces cell proliferation and promotes differentiation( Reference Bartram, Scheppach and Schmid 14 ). Animal experiments have shown that butyrate may reduce colorectal carcinogenesis by enhancing the apoptotic response to methylating carcinogens( Reference Clarke, Young and Topping 15 , Reference Le Leu, Brown and Hu 16 ).
Increasing large-bowel butyrate supply has the potential to improve colonic function and lower disease risk. RS is thought to be particularly effective in this regard as its fermentation generally favours butyrate production. Red meat and fibre (including RS) are generally consumed together as components of foods. Our animal studies have shown that dietary RS is able to oppose colonocyte DNA strand breaks, telomere shortening and pro-mutagenic DNA adduct formation in rodents fed red meat( Reference Winter, Nyskohus and Young 8 , Reference Toden, Bird and Topping 17 , Reference O'Callaghan, Toden and Bird 18 ). This protective effect correlated most closely with large-bowel butyrate levels, supporting a role for fermentation in risk modification. Acylated starches (classified as RS4, chemically modified), in which the acyl group is linked to the starch framework by an ester bond, can deliver specific SCFA to the large bowel where bacterial esterases release the SCFA. Ingestion of butyrylated high-amylose maize starch increases colonic butyrate levels in animals( Reference Clarke, Topping and Bird 19 ) and humans( Reference Clarke, Topping and Christophersen 20 ). Accordingly, the present study was carried out in healthy individuals to determine whether high red meat consumption generated O6MeG adducts in rectal epithelial cells, and whether concurrent consumption of high red meat and butyrylated high-amylose maize starch opposed this effect (primary aim). We also investigated the effects of these diets on other indices of colonic health including rectal proliferation, colonic fermentation products and microbiota composition, as these might participate in the generation of adducts.
Methods
Study design and participants
The present study was conducted as a double-blind, randomised cross-over trial consisting of two intervention periods of 4 weeks each, preceded by a 4-week run-in (baseline) and separated by a 4-week (washout) period (Fig. 1). A group of twenty-three healthy volunteers participated in the trial. Exclusion criteria included evidence of active mucosal bowel disease, intolerance to high-fibre foods or any perceived contraindication to consumption of the test products. At enrolment, all participants showed no active bowel disease. During the entry (baseline) period, participants consumed their habitual diets. For the interventions, they were allocated randomly to a high red meat (HRM) diet or to a HRM diet supplemented with 40 g/d of butyrylated high-amylose maize starch (HRM+HAMSB diet). During the HRM intervention, participants consumed 300 g/d (raw weight) of cooked lean red meat that was supplied frozen in 100 g packs of lean mince, beef strips or lamb strips, with three packs to be consumed each day. During the HRM+HAMSB intervention, participants were also required to consume a total of two pre-packed 20 g sachets of HAMSB daily, one in the morning and one in the evening by mixing the powder into 250 ml of reduced-fat milk or orange juice. HAMSB was manufactured by Ingredion, whereby 23 % of each glucopyranosyl unit in high-amylose maize starch contained a butyrate molecule (degree of substitution 0·23) and was of the same batch as the product used previously( Reference Conlon, Kerr and McSweeney 21 ). Participants on the HRM arm of the study were asked to consume 250 ml of reduced-fat milk or orange juice per d to match the HRM+HAMSB intervention. During the intervention periods, participants reduced their intake of their habitual diet to accommodate the extra 300 g red meat. Participants were instructed to maintain their usual diet during the study but to avoid consuming high levels of protein or fibre, or probiotic supplements, except those prescribed for the study. Participants were also asked to avoid consuming, or record the use of, any medication that could interfere with bowel function (including antibiotics). Participants were monitored by a trial nurse (weekly) and dietitian (at the end of each 4-week period) to ensure that diet and intervention guidelines were followed, and weight was kept stable. Details of medical history and medications, weight, bowel health and symptoms, and adverse events were collected by the trial nurse throughout the study. Composition of the participants' diets and compliance with the interventions was assessed using weighed food diaries that were completed by the participants at the end of each 4-week dietary period, 3 d before each clinic visit. Food diaries were entered into FoodWorks Professional 7 Nutrition Calculation software (Xyris Software) by a dietitian, to calculate energy and macronutrient intake based on Australian food composition tables and food manufacturers' data. The present study was approved by the Flinders Clinical Research Ethics Committee (reference no. 155/09; Flinders Medical Centre, Bedford Park, SA, Australia), and all volunteers gave written informed consent. The present trial was registered in the Australian New Zealand Clinical Trials Registry as ACTRN12609000306213 (http://www.anzctr.org.au).

Fig. 1 Overview of the randomised cross-over intervention study design. HRM, high red meat; HAMSB, butyrylated high-amylose maize starch.
Sample collection
Stool and rectal pinch biopsy samples were obtained at the completion of the 4-week entry period (baseline) and at the end of each intervention period. A complete faecal collection was made by all participants for the last 48 h of each dietary period, and the samples were stored in portable freezers ( − 20°C). At each visit to the Flinders Medical Centre clinic, an experienced gastroenterologist collected four rectal mucosal biopsies using alligator forceps through a 25 cm rigid sigmoidoscope; this procedure was performed without bowel preparation or prior dietary restriction. Biopsies were formalin-fixed and dehydrated through gradient alcohol and xylene before being embedded in paraffin wax.
Stool analyses
Faecal samples were thawed at 4°C, pooled, homogenised, and then subsampled for analysis. For the determination of SCFA, weighed portions were diluted at 1:3 (w/w) with deionised water containing 1·68 mmol heptanoic acid/l as an internal standard (Sigma Chemical Co.), and processed for SCFA analysis using GC as described previously( Reference Winter, Nyskohus and Young 8 ). Total SCFA concentration was calculated as the sum of acetic, propionic, butyric, isobutyric, caproic, isovaleric and valeric acid concentrations. Total branched-chain fatty acids concentration was calculated as the sum of isobutyric and isovaleric acid concentrations. Phenol and p-cresols were measured in the faeces by using vacuum microdistillation and HPLC( Reference King, May and Davies 22 ). Faecal NH3 concentration was measured by using the indophenol blue method( Reference Chaney and Marbach 23 ). Aqueous extracts of the faeces were prepared by diluting 1 g faeces with 4 ml of distilled water, homogenised and centrifuged (4500 rpm, 4°C)( Reference Hughes, Cross and Pollock 24 ), and total apparent NOC were measured by chemical denitrosation with HBr and chemiluminescence detection of the released nitric oxide using a thermal energy analyser (TEA)( Reference Bingham, Pignatelli and Pollock 25 , Reference Russell, Gratz and Duncan 26 ). Concentrations were calculated by comparing the TEA response of a faecal water sample with the response of an N-nitrosodipropylamine standard (16·6 μg/ml), and values were expressed as total apparent NOC (ng/ml)( Reference Holtrop, Johnstone and Fyfe 27 ).
Rectal biopsy analysis
The quantification of the O6MeG adduct load was performed using an immunohistochemical detection method( Reference Winter, Nyskohus and Young 8 ). The immunohistochemical measurement of O6MeG adducts has been previously used for many years mainly in different animal species( Reference Winter, Nyskohus and Young 8 , Reference Zaidi, Potten and Margison 28 – Reference Nyskohus, Watson and Margison 32 ); however, this is the first time it has been applied to human colonic crypts. The specificity of the monoclonal antibody has been validated by RIA( Reference Seiler, Kamino and Emura 33 ). In brief, rectal biopsies were embedded in paraffin and sectioned at 4 μm, and their O6MeG adduct load was quantified using an anti-O6MeG antibody (Squarix Biotechnology); this antibody is listed as being specific for human tissue. Antigen retrieval (10 mm-citrate buffer) was performed, followed by RNase treatment (20 μl RNase A (10 mg/ml), Thermo Fisher Scientific; 5 μl RNase T (10 units/ml), Thermo Fisher; 100 μl PBS (pH 7·4) and stopped with a 5 min treatment with NaCl solution (140 mm)). DNA unwinding was achieved using alkali treatment (70 mm-NaOH/140 mm-NaCl, 1·5 ml) before applying Special Block A (Covance Laboratories) for 30 min. The O6MeG antibody (1:1000) was applied to the slides overnight at room temperature, followed by Special Block B (Covance Laboratories), before applying poly-horseradish peroxidase (HRP) anti-mouse IgG. Sections were counterstained with haematoxylin, and chromogen 3,3′-diaminobenzidine tetrahydrochloride (DAB) was used to visualise positive O6MeG staining. All slides were independently and randomly coded before quantification of nuclear staining for O6MeG with a computer image analysis protocol( Reference Winter, Nyskohus and Young 8 ). Overall, twenty appropriate crypts were visualised using an Olympus Micropublisher 3.3 RTV camera and Olysia Bio-report software (Olympus). Camera and microscope settings were calibrated before each image to ensure analytical consistency. To identify a linear path through a single row of nuclei along the crypt axis for all images taken, image analysis software developed by the CSIRO Mathematics Informatics and Statistics division, ‘Imview’ and ‘R for Windows’ 2.1.0, was used. Raw colour (red, green and blue), luminescence (L), normalised colour values (r= ¼ red/L, g= ¼ green/L and b= ¼ blue/L) and colour ratio (RoB = ¼ r/b) data points were calculated for each pixel along the length of the linear path. The number of cells within each half crypt was counted, and the calculated RoB ratio was then averaged for each nucleus within individual crypts. Total O6MeG values/crypt were achieved by summation of the ratio value for each nucleus along the crypt axis. Representative sections of one individual from each treatment group showing the immunohistochemical staining are shown in Fig. 2. Proliferation status of cells in the rectal crypts was determined by standard immunohistochemical techniques using the proliferating cell nuclear antigen (PCNA) antibody (PC10), as reported previously( Reference Le Leu, Hu and Brown 34 , Reference Winter, Young and Hu 35 ). Slides were visualised by brown nuclear staining, and assessed as the number of Ki-67-positive cells/crypt.

Fig. 2 Light microscope images (20 × optical zoom) of human rectal crypts showing O6-methyl-2-deoxyguanosine staining intensity from the baseline and after the 4-week intervention phase selected from participant #20. Images showing the sections (a) at the end of the high red meat (HRM) baseline, (b) at the end of the 4-week HRM treatment, (c) at the end of the HRM+butyrylated high-amylose maize starch (HAMSB) baseline and (d) at the end of the 4-week HRM+HAMSB treatment.
Molecular microbiology
Extraction of DNA from stool samples and subsequent quantitative real-time PCR (qPCR) was performed and analysed according to the method used by Christophersen and colleagues( Reference Christophersen, Morrison and Conlon 36 ). In brief, DNA was extracted using a repeat bead beating and column clean-up method, and qPCR assays amplified the 16S ribosomal RNA (rRNA) gene (or in the case of sulphate-reducing bacteria the adenosine-5′-phosphosulfate reductase (aps) gene) using primers that targeted bacterial species or groups of interest. The primer pairs and their amplification conditions are listed in online Supplementary Table S5. Data are expressed as absolute abundances and as a proportion of total bacteria. Bacterial targets were chosen for their relevance to gut health. In other words, we selected key species (e.g. F. prausnitzii) or groups of bacteria (e.g. C. coccoides group) that were responsible for the production of butyrate following fermentation of complex carbohydrates, particularly RS, by bacteria such Ruminococcus bromii. P. distasonis was chosen because of its potential role in the cleavage of butyrate from the butyrylated RS used in the study. We also examined changes in some potentially enteropathogenic species (e.g. E. coli), as well as in general groups such as Lactobacillus and Bifidobacterium that are regarded as markers of bowel health by many. A range of other bacteria such as those implicated in gut mucus barrier turnover and inflammatory bowel disease, such as A. muciniphila, were also targeted. Sulphate-reducing bacteria were included to determine whether the production of toxic hydrogen sulphide could contribute to large-bowel DNA adduct formation in response to red meat treatment.
Statistical methods
All statistical analyses were performed with scripts in R, version 3.0.1, using the R statistical package( 37 ). Analyses were carried out using a linear mixed-effects model, with subjects as the random effect, on either base-10 logarithm-transformed data, where necessary, or on untransformed data for each of the variables.
Initial analyses were carried out using the two periods of the trial, including the baseline and washout periods. However, data analyses of the initial study showed that some of the response variables had carry-over effects, including the primary end-point O6MeG, epithelial proliferation, certain bacterial species but not SCFA (see the online Supplementary material for a full study dataset). This was indicated by a significant difference between the baseline level and the washout level of the response variable or a significant interaction between the week of diet consumption and the response variable. Therefore, the data analyses used in the present study were only those of the first period of the study (i.e. only measurement weeks 0 and 4). As a result, the analysis reported herein was carried out using only the baseline and the first-period data. The comparison between the groups in the first period of the trial was carried out using a linear mixed-effects model, testing for changes from the baseline and a difference between the treatments. For Tables 1–4, dietary intake, stool biochemistry, bacterial abundance (percentage of total bacteria) and rectal biology data for the first period of the trial are expressed as means with their standard errors of the mean for both groups (HRM and HRM+HAMSB), together with the increment and percentage change for each group. For each of these means, the significance of the change from the baseline is indicated. The final column of each table gives the P value for the difference between the two treatments at week 4, and these were tested using either the original data or the log 10-transformed data as appropriate.
Table 1 Dietary intake of the study participants during each diet period, based on 3 d weighed food records (Mean values with their standard errors; percentages)

HRM, high red meat; HAMSB, butyrylated high-amylose maize starch.
Mean value was significantly different from that at baseline: * P< 0·05, ** P< 0·01, *** P< 0·001 (linear mixed-effects model).
† HAMSB supplement contains 88 % total carbohydrate, approximately 20 % dietary fibre, 10 % moisture, < 1 % total fat and < 0·75 % protein.
‡ P value was obtained for treatment difference at week 4 (linear mixed-effects model).
§ P< 0·05.
Table 2 Effect of the dietary interventions in the first period on rectal biology (Mean values with their standard errors; percentages)
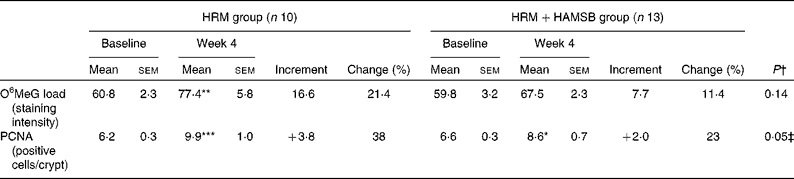
HRM, high red meat; HAMSB, butyrylated high-amylose maize starch; O6MeG, O6-methyl-2-deoxyguanosine; PCNA, proliferating cell nuclear antigen.
Mean value was significantly different from that at baseline: * P< 0·05, ** P< 0·01, *** P< 0·001 (linear mixed-effects model).
† P value was obtained for treatment difference at week 4 (linear mixed-effects model).
‡ P< 0·05.
Table 3 Effect of the dietary interventions in the first period on stool biochemistry (Mean values with their standard errors; percentages)

HRM, high red meat; HAMSB, butyrylated high-amylose maize starch; BCFA, branched-chain fatty acids; NOC, N-nitroso compounds.
Mean value was significantly different from that at baseline: * P< 0·05, ** P< 0·01, *** P< 0·001 (linear mixed-effects model).
† P value was obtained for treatment difference at week 4 (linear mixed-effects model).
‡ P< 0·05.
Table 4 Abundances of species and groups of bacteria (per g of stool and as a percentage of total bacteria)† (Mean values with their standard errors; percentages)

HRM, high red meat; HAMSB, butyrylated high-amylose maize starch.
Mean value was significantly different from that at baseline: * P< 0·05, ** P< 0·01, *** P< 0·001 (linear mixed-effects model).
† As enumerated using quantitative real-time PCR and showing significant changes in response to the dietary treatments.
‡ P value was obtained for treatment difference at week 4 (linear mixed-effects model).
§ P< 0·05, §§ P< 0·01, §§§ P< 0·001.
The effects on the overall composition of the gut microbiota were analysed by combining all qPCR assays and performing a permutational-based multivariate analysis. Data were log-transformed before producing a resemblance matrix using Euclidean distance. Differences between the interventions were tested on first-period cross-sectional comparison only using Permanova+ version 1.06 (PRIMER-E). A P value < 0·05 was considered significant.
Results
Study comparisons for the whole dataset
Initial analyses were carried out using the two periods of the trial, including baseline and washout periods. However, data analyses of the initial study showed that some of the response variables had carry-over effects, including the primary end-point O6MeG, epithelial proliferation, certain bacterial species but not SCFA (see online Supplementary material for a full study dataset). There was a significant increase in the rectal crypt O6MeG adduct load when the participants consumed the HRM diet first compared with all the other intervention stages (P< 0·01; see online Supplementary Fig. S2(A)); however, when the participants consumed the HRM+HAMSB diet as the first intervention, there was no change in the O6MeG adduct load with the subsequent consumption of HRM (see online Supplementary Fig. S2(B)). There was a significant effect of treatment and treatment order on PCNA-positive cells/crypt (see online Supplementary Fig. S3(A) and (B)). For the participants on the HRM or HRM+HAMSB diet as their first intervention, the PCNA-positive cells significantly increased (P< 0·001). For those on the HRM or HRM+HAMSB diet (received as their first intervention), the positive cells significantly decreased after consuming their final treatment compared with the first treatment. Participants who consumed the HRM diet as the first treatment had significantly higher PCNA-positive cells/crypt compared with those who consumed the HRM diet as the second treatment (P< 0·001). We also observed that numbers of some bacteria in the washout phase were significantly different from those during the entry period or the dietary interventions (see online Supplementary Tables S3 and S4). The analysis revealed a diet-order effect on the microbiota composition. When the volunteers consumed the HRM+HAMSB diet as the first intervention, their microbiota composition significantly differed from that of the entry levels (P= 0·02), as well as from that of the subsequent HRM intervention (P= 0·02) and the washout levels (P= 0·005) in the same volunteers. Furthermore, the microbiota composition of these volunteers consuming the HRM+HAMSB diet first was also significantly different from that of those consuming HRM first (P= 0·01). However, when the volunteers consumed the HRM diet first, the microbial composition during the subsequent consumption of the HRM+HAMSB diet was only different from that of the washout levels (P= 0·02).
Study comparisons for the first period
The results arising from the respective baseline periods and the first arm of the dietary intervention (i.e. at week 4 of the intervention) are described in detail below, as the results of the second arm (cross-over) showed carry-over effects for O6MeG, epithelial proliferation and certain bacterial species. The study was still adequately powered based on the primary end-point ‘O6MeG’. Calculations using a two-tailed t test with a power of 80 % with means of 60·8 and 77·4 and a standard deviation of 15 showed that group sizes of five were adequate to detect a difference between the baseline and the HRM intervention.
Demographic data, participant characteristics and dietary intake
Recruitment commenced in July 2009, with each participant followed up for the 4-month duration of the interventions. Data collection was completed by September 2010. A total of twenty-five participants were assigned randomly, with twelve allocated to the HRM dietary intervention first and thirteen allocated to the HRM+HAMSB dietary intervention first. However, two participants withdrew before the commencement of the intervention diets; one due to unrelated medical problems and the other due to intolerance of the first rectal biopsy. Approximately one-third of the participants on the trial diets reported increased flatulence. Of the volunteers, ten (seven males and three females; mean age 62·1 (sem 1·8) years and mean body weight 79·8 (sem 5·6) kg) completed the HRM intervention as the first diet period, while thirteen (ten males and three females; age 62·7 (sem 1·7) years and body weight 82·4 (sem 3·5) kg) completed the HRM+HAMSB intervention first.
Participants maintained consistent body weight, with mean weights of 77·1 (sem 6·4) and 82·8 (sem 3·3) kg after the HRM and HRM+HAMSB interventions, respectively.
There was no difference between the diets for reported intake of energy, total and saturated fat, total carbohydrates and sugar, starch, alcohol or total Fe intake (Table 1). Compared with their respective baseline levels, protein intake was significantly increased by the HRM (P< 0·05) and HRM+HAMSB (P< 0·01) interventions. Fibre intake was decreased in the HRM group at week 4 compared with its baseline level (P< 0·01). Fibre intake was significantly lower in the HRM group than in the HRM+HAMSB group after 4 weeks (P< 0·01). Fe intake from meat was significantly higher for the HRM and HRM+HAMSB interventions at week 4 compared with their respective baseline levels (P< 0·001).
Rectal epithelial measures
The O6MeG adduct load was increased at week 4 in the HRM group compared with its baseline level (P< 0·01); however, the addition of HAMSB to the HRM diet prevented this increase (Table 2). Relative to their respective baseline levels, the number of PCNA-labelled cells in the rectal epithelium increased for both the HRM (P< 0·001) and HRM+HAMSB (P< 0·05) groups; however, the number was lower following the HRM+HAMSB intervention than the HRM intervention (P< 0·05; Table 2).
Stool biochemistry
The results of the stool analyses are presented in Table 3. Stool output and pH did not differ significantly between the treatments. Stool excretion of acetate, propionate, butyrate and total SCFA was higher in the HRM+HAMSB group at 4 weeks compared with its baseline level (P< 0·05) as was the stool concentrations of acetate, propionate and total SCFA (P< 0·05). Faecal p-cresol concentration was lower in the HRM+HAMSB group at 4 weeks compared with its baseline level (P< 0·01) and the HRM group (P< 0·05). Branched-chain fatty acids, phenols, NH3 and NOC were unaffected by the treatment.
Stool bacteria
Significant changes in stool bacteria in response to the diet are presented in Table 4. When the numbers of bacteria/g of stool were examined, the HRM+HAMSB group at 4 weeks elicited an increase in the number of Parabacteroides distasonis relative to its baseline level (P< 0·0001) and the HRM group (P< 0·001). Compared with its baseline level, HRM+HAMSB consumption increased the numbers of Lactobacillus spp. (P< 0·01), the Clostridium coccoides group (P< 0·05), the Clostridium leptum group (P< 0·05) and Ruminococcus bromii (P< 0·05), but lowered the numbers of Ruminococcus torques (P< 0·05). When bacterial numbers were expressed as a percentage of total bacteria, the proportion of P. distasonis was increased by the consumption of the HRM+HAMSB diet at 4 weeks compared with its baseline level and the HRM group at 4 weeks (both P< 0·0001). Lower proportions of Ruminococcus gnavus (P< 0·01), R. torques (P< 0·01) and E. coli (P< 0·01) were evident in the HRM+HAMSB group at 4 weeks compared with its baseline level. The HRM+HAMSB group also had lower proportions of R. torques (P< 0·05) and E. coli (P< 0·05) than the HRM group at 4 weeks. When the results of all qPCR assays were combined and then analysed statistically to gain an indication of the impacts of the treatments on microbial diversity, it was found that the microbial diversity during the HRM+HAMSB intervention was different from that at baseline (P< 0·05) and during the HRM intervention (P< 0·01); however, the composition at baseline and the HRM intervention did not differ.
Discussion
Previously, we reported that feeding a diet rich in red meat to rodents can increase the level of the pro-mutagenic DNA adduct (O6MeG) in the colon, whereas co-consumption of a fermentable carbohydrate can reduce this effect( Reference Winter, Nyskohus and Young 8 ). We have now shown that when free-living healthy human subjects consumed their normal habitual diet containing at least an additional 300 g red meat over a 4-week period, there was increased formation of the O6MeG adduct in the rectal epithelium. This increase in adduct formation might account, in part, for the increased risk of CRC associated with consuming high levels of red meat.
Studies in rodents have shown a positive correlation between cumulative O6MeG levels and tumour load( Reference Povey 38 ). This association is also supported, in humans, by the prevalence of higher O6MeG levels in DNA isolated from the distal region of the colon, where most sporadic CRC occurs( Reference Povey, Hall and Badawi 39 ). The present study is the first to report on the effect of feeding a HRM diet to human subjects on the most predominant alkyl-induced DNA adduct O6MeG in the rectal epithelial tissue. In a randomised cross-over study comparing HRM, vegetarian and HRM/high-fibre diets, an increase in O6-carboxymethylguanine adduct levels was observed in exfoliated colonic epithelial cells isolated from the faeces of healthy volunteers consuming a HRM diet( Reference Lewin, Bailey and Bandaletova 7 ). However, the relevance of DNA adducts in exfoliated cells to the in situ epithelial adduct load is unclear. Our findings show that such adducts do form in cells residing within the crypt and, thus, have the potential to form mutated clones that might progress to cancer.
The present study also confirms that dietary fermentable carbohydrate in the form of HAMSB can protect against red meat-induced colorectal DNA lesions in humans, and is consistent with epidemiological evidence that dietary fibre consumption reduces the risk of CRC. The present study and our previous work in rodents( Reference Winter, Nyskohus and Young 8 , Reference O'Callaghan, Toden and Bird 18 , Reference Conlon, Kerr and McSweeney 21 , Reference Toden, Bird and Topping 40 ) all point towards SCFA, particularly butyrate, to be the key mediators in preventing meat-induced DNA adducts and DNA strand breaks in colonic mucosa. Butyrate is a preferred metabolic substrate for colonocytes, and this SCFA has strong anti-tumorigenic properties in vivo and in vitro ( Reference Clarke, Young and Topping 15 , Reference Le Leu, Hu and Young 41 ). In the present study, ingestion of HAMSB in conjunction with HRM was also able to favourably influence the colonic luminal environment, as evidenced by increased levels of SCFA and a reduction in the potentially toxic protein fermentation product p-cresol. This elevation of faecal butyrate with HAMSB confirms previous studies in human subjects( Reference Clarke, Topping and Christophersen 20 , Reference Clarke, Bird and Topping 42 ), and has the potential to improve colonic health and offer protection against CRC. Although consumption of a blend of types 2 and 3 RS in a recent human trial of hereditary CRC failed to reduce tumour incidence( Reference Mathers, Movahedi and Macrae 43 ), the relatively low daily intake of RS used in that study may have been insufficient to increase SCFA levels in stool (which were not measured). At least 20 g of RS/d may be needed to increase stool levels of SCFA( Reference Topping, Bird and Young 44 , Reference Worthley, Le Leu and Whitehall 45 ).
A reasonable explanation for the increase in O6MeG adducts with the HRM intervention is dietary haem. Haem is abundant in red meat, the majority of which derived from the diet passes into the large bowel( Reference Young, Rose and St John 46 ). We recently identified dietary haem as an agent that can increase O6MeG adducts in the colon of mice( Reference Winter, Young and Hu 35 ). Haem Fe-rich meat has also been shown to increase alkylated DNA adducts in an in vitro digestion system( Reference Vanden Bussche, Hemeryck and Van Hecke 47 ). Dietary haem may also increase the production of reactive oxygen species, causing cellular toxicity and pro-mutagenic lesions( Reference Glei, Klenow and Sauer 48 , Reference Knobel, Weise and Glei 49 ). Other factors, such as bile acids( Reference Ridlon, Kang and Hylemon 50 ), could also contribute to adduct formation and DNA damage more broadly. Haem may also be responsible for the increased rectal cell proliferation in response to HRM and HRM+HAMSB consumption, as evidenced by more PCNA-labelled cells/crypt. Haem is associated with increased epithelial proliferation in the colon of rodents, and can injure the colonic surface epithelium by generating cytotoxic and oxidative stress( Reference Rijnierse A and de Wit 6 , Reference de Vogel, Jonker-Termont and van Lieshout 51 ).
In the present study, we anticipated that NOC would increase in the stool of the participants consuming the HRM diet, and that this would explain a higher O6MeG adduct load. High dietary haem and red meat have previously been associated with increased luminal NOC in humans( Reference Hughes, Cross and Pollock 24 , Reference Holtrop, Johnstone and Fyfe 27 , Reference Lunn, Kuhnle and Mai 52 ). A dose–response relationship has been described between red meat intake and faecal NOC: low faecal NOC (374 μg/kg) at low red meat intake (60 g/d) and a 4- or 5-fold increase in faecal NOC with increased red meat intake of 240 and 420 g/d, respectively( Reference Hughes, Cross and Pollock 24 ). Lewin et al. ( Reference Lewin, Bailey and Bandaletova 7 ) also observed an increase in faecal NOC in volunteers fed 420 g/d of red meat in comparison with a vegetarian diet, and suggested that NOC are important genotoxins involved in the generation of alkyl adducts( Reference Lewin, Bailey and Bandaletova 7 ). Although we observed an increase in the O6MeG adduct load with HRM intake (300 g/d), we are unable to completely explain the lack of the effect of HRM on faecal NOC. One possible explanation is that the other studies( Reference Lewin, Bailey and Bandaletova 7 , Reference Hughes, Cross and Pollock 24 ) had a very high level of control with the meals being consumed in an experimental facility. In the present study, we only controlled for the amount of meat in the diet so that other factors in the diet may account for the discrepancy.
There is growing recognition of both the importance of the large-bowel microbiota in human health and the strong role of diet in modulating its composition and metabolic activities. Using a suite of qPCR assays that targeted a range of bacteria important to gut health, we demonstrated significant shifts in the composition of the gut microbiota in response to the HRM+HAMSB, but not HRM, intervention. Our observation of an increase in the stool numbers of the C. leptum group and R. bromii, a member of the C. leptum group, in response to the consumption of RS as HAMSB is consistent with the effects previously observed in human subjects and with the central role that this bacterium appears to have in RS degradation( Reference Martinez, Kim and Duffy 53 – Reference Abell, Cooke and Bennett 55 ). This increase provides further evidence (additional to the observed increase in stool SCFA levels) that HAMSB was being consumed by the participants and was reaching the large bowel where it was available for fermentation. The numbers of P. distasonis were also increased by the HAMSB treatment, which is also consistent with changes in humans following consumption of butyrylated RS( Reference Clarke, Topping and Christophersen 20 ). HAMSB is a chemically modified RS, and the forms of RS (classified as RS4) have been shown to be more likely to stimulate the growth of P. distasonis ( Reference Martinez, Kim and Duffy 53 ). In line with the HAMSB treatment, stool excretion of butyrate increased, primarily from the release of the bound butyrate; however, the higher number of bacteria in the C. coccoides group could also have contributed as a number of butyrate producers are classified in that group.
When the numbers of bacteria were expressed as a proportion of total bacteria, the addition of HAMSB to the diet also lowered the numbers of E. coli (a species with enteropathogenic variants and potential), R. gnavus and R. torques (numbers of which are high in the mucosa of some individuals with inflammatory bowel disease)( Reference Png, Linden and Gilshenan 56 ), supporting the potential of HAMSB to promote gut health. We did not observe any clear indication of the effects of HRM on the composition of the gut microbiota. However, the range of bacteria that we targeted is limited, and a more detailed analysis of populations may reveal changes. This may give an insight into the mechanisms of HRM-induced adduct formation and reasons for the associated increased risk of CRC. Furthermore, in vitro experiments have demonstrated that the formation of alkylated DNA adducts appear to depend on the microbial composition( Reference Vanden Bussche, Hemeryck and Van Hecke 47 ). The changes that we have observed support the idea that the increases in stool SCFA levels and the associated protection against dietary HRM-mediated colorectal tissue damage that have occurred in response to dietary RS treatment are at least partly mediated by the gut microbiota, through both cleavage of the esterified butyrate and fermentation of the RS substrate. Part of the protective effect may also be attributable to reducing numbers and activities of bacteria with potential for harm.
A limitation of the present study was that the randomised cross-over design resulted in a period effect for the primary end-point ‘O6MeG adducts’. Participants allocated the HRM+HAMSB intervention in the first period did not have a HRM-induced increase in O6MeG adducts during the subsequent period. This contrasted with the increase in adduct formation that occurred when HRM was consumed first. This suggests that the consumption of HAMSB is able to protect against the damage caused by the HRM diet. We have presented in detail the analyses of the data from the first period (results incorporating both treatment periods are supplied in the online Supplementary material). Presentation of the data from the first arm reduces the statistical power of the study compared with that of the full cross-over. However, the primary end-point ‘O6MeG’ was still adequately powered, and the effect of HAMSB on SCFA (especially butyrate) is magnified. A further limitation of the study was that the right-sided colonic mucosa could not be evaluated, as biopsies were only taken from the rectum. This limits the application of the results to proximal colorectal carcinogenesis, particularly as genotypic differences between proximal and distal cancers exist( Reference Iacopetta 57 ). More invasive studies are warranted to investigate whether the same effects observed in the rectum occur in other regions of the large bowel.
In summary, our findings show that high dietary red meat intake has detrimental effects on the colorectum by increasing pro-mutagenic DNA adducts and epithelial cell proliferation. Conversely, increasing luminal butyrate levels with HAMSB prevented the accumulation of O6MeG adducts. These findings might explain the increased risk for CRC associated with HRM consumption, and could point to a beneficial effect of butyrate-generating RS.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0007114515001750
Acknowledgements
The authors thank the volunteers who took part in the study, and the clinical trial nurses Libby Bambacas and Jane Upton for their help in co-ordinating the volunteers and sample collection. The authors thank Penny Taylor for providing the volunteers with dietitian advice and preparation of the diet information booklet used for the study. The authors thank Michelle Vuaran and Jennifer Giles for their assistance with the microbiology analysis and Jenny McInerney and Sylvia Usher for assistance with the analysis of fermentation products. The authors are also indebted to Dr Julie Clarke for arranging the supply of butyrylated starch and for its preclinical evaluation.
The present study was financially supported by the National Health and Medical Research Council of Australia (project no. 535079) and the Commonwealth Scientific and Industrial Research Organisation (CSIRO) Preventative Health Flagship, Australia.
The authors' responsibilities were as follows: R. K. L., G. P. Y., M. A. C., Y. H., D. L. T. and A. R. B. designed and conducted the study; R. B. M. and C. T. C. performed statistical analyses; J. M. W. and K. J. H. performed analytical analysis; S. W. G. coordinated NOC analysis; K. J. H. performed analysis of the dietary records; C. T. C. and M. A. C. performed bacterial analysis and interpretation; R. K. L., M. A. C., G. P. Y., D. L. T., Y. H., J. M. W., A. R. B., S. W. G., C. T. C. and K. J. H writing and manuscript preparation. All authors read and approved the final manuscript.
The authors declare that they have no conflicts of interest.









 Open access
Open access