Introduction
Human schistosomiasis is one of the most prominent neglected tropical diseases (NTDs) in Africa (Hotez & Kamath, Reference Hotez and Kamath2009). The disease is caused by water-borne, snail-transmitted trematode parasites of the genus Schistosoma, and is widely distributed in 78 countries with about 206 million cases and 2.5 million disability adjusted life years (World Health Organization, 2022). There are six species of the genus Schistosoma infecting humans worldwide, with four common species in Africa: Schistosoma haematobium, Schistosoma mansoni, Schistosoma guineensis and Schistosoma intercalatum. Schistosoma haematobium is responsible for most of the morbidity in Africa, with the adult parasite inhabiting the vesicular and pelvic venous plexus of the bladder and causing urogenital schistosomiasis (World Health Organization, 2021). The pathologies associated with this species is dependent on the severity of infection, migration of the worms and inflammatory responses to the presence of the eggs (Centers for Disease Control, 2021; World Health Organization, 2021).
Largely, schistosomiasis is a focal disease (Global Schistosomiasis Alliance, 2020), which thrives in rural and marginalized urban populations that share proximities with surface waterbodies containing the appropriate susceptible species of freshwater snail vectors, and are characterized by inadequate or poor access to water, sanitation and hygiene (WASH) facilities (Olamiju et al., Reference Olamiju, Nebe and Mogaji2022). In such areas, contact activities with surface waterbodies ranges from bathing, washing of clothes, swimming and playing are not uncommon (Olamiju et al., Reference Olamiju, Nebe and Mogaji2022). These activities support the transmission cycle of the parasites, with infestation of water bodies through open urination or defecation by an infected resident, and subsequent infection of other residents through other domestic contact purposes. Control and elimination for schistosomiasis have therefore focused on mass administration of praziquantel to children between ages 5 and 15 years in endemic communities (World Health Organization, 2022), with possible complementary provision of WASH interventions to promote behavioural change (Grimes et al., Reference Grimes, Croll, Harrison, Utzinger, Freeman and Templeton2015). However, in endemic communities, with infested surface water bodies, where both humans and livestock interact, there are growing concerns on the hybridization of closely related species of humans (S. haematobium) and cattle (Schistosoma bovis) (Léger et al., Reference Léger, Garba, Hamidou, Webster, Pennance, Rollinson and Webster2016). Hybrids are commonly identified based on discordance between nuclear and mitochondrial markers (Onyekwere et al., Reference Onyekwere, Rey, Allienne, Nwanchor, Alo, Uwa and Boissier2022). The emergence of hybrid lineages has raised significant concerns for schistosomiasis control and elimination effort (Léger & Webster, Reference Léger and Webster2017), although the extent to which hybridization is actually occurring at present is subject to debate (Platt et al., Reference Platt, McDew-White and Le Clec'h2019). We therefore hypothesize that the presence of atypical characteristics (morphology and morphotype) of Schistosoma eggs may serve as a potential indicator in the detection of possible hybridization cases, since different egg morphotypes have been observed among patients infected with S. haematobium × S. bovis hybrids (Moné et al., Reference Moné, Minguez, Ibikounlé, Allienne, Massougbodji and Mouahid2012, Reference Moné, Holtfreter and Allienne2015; Soentjens et al., Reference Soentjens, Cnops, Huyse, Yansouni, De Vos, Bottieau, Clerinx and Van Esbroeck2016; Boon et al., Reference Boon, Fannes, Rombouts, Polman, Volckaert and Huyse2017).
Nigeria has the highest burden of schistosomiasis in Africa (Hotez & Kamath, Reference Hotez and Kamath2009), with the disease being endemic across all 36 States in the country (Federal Ministry of Health, 2019). Around certain transmission foci, precisely, communities situated along the banks of Oyan-dam in the south-western part of the country, prevalence can reach as high as 90% (Akinwale et al., Reference Akinwale, Ajayi, Akande, Gyang, Adeleke, Adeneye, Adebayo and Dike2010). The communities (Abule-Titun, Apojola, Ibaro and Imala-Odo) have remained highly endemic since 1991 despite ongoing interventions (Ekpo et al., Reference Ekpo, Odeyemi, Sam-Wobo, Onunkwor, Mogaji, Oluwole, Abdussalam and Stothard2017). Predominant water contact activities such as fishing, farming, bathing, swimming, drinking, washing clothes or kitchen utensils and fetching of water from infested surface water bodies are common (Akinwale et al., Reference Akinwale, Ajayi, Akande, Gyang, Adeleke, Adeneye, Adebayo and Dike2010; Ekpo et al., Reference Ekpo, Odeyemi, Sam-Wobo, Onunkwor, Mogaji, Oluwole, Abdussalam and Stothard2017). Furthermore, livestock farming is one of the most common occupations of the populace, and this allows interaction between cattle with humans along the banks of the dam that surrounds these communities (fig. 1). Since S. bovis has been previously reported as the predominant livestock species in the country (Hambali et al., Reference Hambali, Adamu, Ahmed, Bokko, Mbaya, Tijjani, Biu, Jesse and Ambali2016; Akande & Alohuntade, Reference Akande and Alohuntade2021), we therefore hypothesize that Schistosoma hybridization and zoonotic transmission may be ongoing in Nigeria in obscurity. This present study therefore aims to characterize the morphology of Schistosoma eggs recovered from urine samples of humans living along the banks of the Oyan River Dam in the south-western part of Nigeria, as part of an ongoing effort towards detection of possible hybridization cases.

Fig. 1. Sharing of common water source by humans and cattle at Apojola community.
Methods
Study communities
This study was conducted in four communities along the shoreline of Oyan River Dam in Ogun State, Nigeria. Ogun State is one of the 36 States in the Federal Republic of Nigeria, located in the south-western region of the country (fig. 2). It covers a total area of 16,409.26 sq. km between latitude 6.2°N and 7.8°N and longitude 3.0°E and 5.0°E. The State has 20 local government areas (LGAs) comprising 236 political wards. The greater proportion of the State lies in the tropical rainforest zone, while the far northern part has features of the Guinea Savannah. In the early 1990s, the Oyan River Dam was established, with its shorelines around four major communities: Imala-Odo and Ibaro-Oyan in Abeokuta North LGA, and Apojola and Abule-Titun in Odeda LGA (fig. 2). These communities have remained endemic for schistosomiasis since 1991 (Ekpo et al., Reference Ekpo, Odeyemi, Sam-Wobo, Onunkwor, Mogaji, Oluwole, Abdussalam and Stothard2017), with predominant occupations being fishing and livestock farming (Akinwale et al., Reference Akinwale, Ajayi, Akande, Gyang, Adeleke, Adeneye, Adebayo and Dike2010). This in addition to poor access to improved WASH facilities have also promoted water contact activities such as bathing, swimming, drinking, washing clothes or kitchen utensils and fetching of water from surface water bodies, and more importantly shared interaction between cattle with humans along the shoreline of the dam that surrounds these communities (fig. 1).

Fig. 2. Map of Ogun State showing the local government areas (LGAs) and study communities and river system.
Sample size determination and recruitment of study participants
The sample size for this study was determined using the formula described by Thrusfield (Reference Thrusfield2005), n s = (1.96/d)2 ⋅ (p(1 − p) where ns is the sample size, p is the existing prevalence in the study area, and d is the degree of accuracy. In determining the sample size, a prevalence of 47% (Ekpo et al., Reference Ekpo, Odeyemi, Sam-Wobo, Onunkwor, Mogaji, Oluwole, Abdussalam and Stothard2017), and a degree of accuracy of 5% was considered at 95% level of confidence. The minimum sample size determined therefore was 383, that is, an average of 96 persons per community. The communities were compact, and invitations to participate were sent to all residents through a local mobilizer. Recruitment and collection of samples was done at a central location and only residents who consented to the study procedures were enrolled into the study.
Sample collection and examination
Urine samples were collected from consenting residents comprising infants and preschoolers (1–4 years), school-aged children (5–15 years) and those above 16–64 years across study communities between October and November, 2019. Age of participants were validated using birth-cards to avoid information bias. Three-hundred and eighty-four samples were collected in dark, sterile 30 ml universal containers and preserved with 70% ethanol. Collections were made between 10.00 and 14.00 h as recommended (Ekpo et al., Reference Ekpo, Laja-Deile, Oluwole, Sam-Wobo and Mafiana2010) and transported to the laboratory in ice boxes. Urine samples were processed using sedimentation techniques, and examined under the microscope for the presence of S. haematobium eggs. A total of 219 (57.0%) of the samples examined were positive for S. haematobium eggs, and separated for subsequent screening using the morphometric method. Participants who were positive with S. haematobium eggs were treated with praziquantel.
Morphometric analysis of S. haematobium eggs
A total of 1984 eggs were recovered from urine sediment of infected participants. Eggs were picked at random and examined using morphometric methods. Microphotographs of the Schistosoma eggs and ova were taken using an AmScope MD130 1.3MP Digital Microscope (United Scope LLC., CA, USA) and the IC Measure™ (The Imaging Source Europe GmbH, Bremen, Germany) computer software was used to measure the total length (including the terminal spine) and the maximum width. The egg length/width ratio was subsequently computed. Qualitative characteristics such as unusual morphology was noted, and the presence or absence of the terminal spine was also recorded. The eggs were classified as typical if they have a round-to-oval shape or atypical if they have spindle/elongated shape (Pitchford, Reference Pitchford1965; Boon et al., Reference Boon, Fannes, Rombouts, Polman, Volckaert and Huyse2017). Atypical schistosomes were identified by their spindle egg-shape (Moné et al., Reference Moné, Holtfreter and Allienne2015; Boon et al., Reference Boon, Fannes, Rombouts, Polman, Volckaert and Huyse2017). A total of 639 (32.2%) eggs were characterized with atypical shape.
Data analysis
Data collected were analysed using SPSS version 23.0 for Windows. Descriptive statistics and differences in proportions were tested using the Chi-square statistics, either for trend or independence, as appropriate. The number of eggs counted was transformed using logarithmic function (log (n + 1)), to normalize the distribution of the residual values for statistical analyses. Morphometric data were exported from the IC Measure™ into Microsoft Excel for analysis. Differences between means between the egg morphotypes across the study communities were tested using an independent sample t-test, and statistical difference was set at 95% confidence interval (P-value < 0.05). The corresponding dataset and other supporting files have been attached as supplementary files.
Results
Demographic characteristics of study participants and infection status
A total of 384 participants were recruited, 198 (51.6%) males and 186 (48.4%) females, between the age group 1–4 years (112; 29.2%), 5–15 years (190; 49.5%) and 16–64 years (82; 21.4%). An overall prevalence of 219 (57.0%) was recorded, with the majority of the infection among the participants from Ibaro-Oyan with a prevalence of 62.4%. Also, the prevalence of infection was higher in the males (31.8%) and in the 5–15 years age group (26.0%) (table 1).
Table 1. Demographic characteristics and infection status of study participants.

Morphotypes of Schistosoma eggs across the study areas
A total of 1984 schistosome eggs from 219 (57.0%) infected individuals were photographed and measured. This comprised 605 (30.5%), 313 (15.8%), 775 (39.1%) and 291 (14.7%) eggs from Imala-Odo, Abule-Titun, Apojola and Ibaro-Oyan, respectively (table 2). Three egg morphotypes were recorded in this study (figs 3–5). The majority of the schistosome eggs were of the typical round-to-oval shape (1345, 67.8%), and atypical elongated or spindle-shape (639, 32.2%). By location, 417 (68.9%), 192 (61.3%), 523 (67.5%) and 213 (73.2%) of the eggs recovered from Imala-Odo, Abule-Titun, Apojola and Ibaro-Oyan were round-to-oval shape, while 188 (31.1%), 121 (38.7%), 252 (32.5%) and 78 (26.8%) were spindle-shaped, respectively (table 2). There was a significant variation in egg morphotypes across the study area (P = 0.017). About 98.7% and 99.2% of the typical and atypical eggs were with spines. There was also a significant variation in the presence of spined eggs across the study area (P = 0.05). However, the majority of the spineless Schistosoma eggs (17, 77%) were of the typical round-to-oval shape, and (5, 23%) with the spindle shape. The typical-shaped but spineless eggs were recovered from Imala-Odo (8, 1.3%), Apojola (5, 0.6%) and Ibaro-Oyan (4, 1.4%), while the spineless spindle-shaped eggs were recovered from Abule-Titun (5, 1.6%). There was also a significant variation in the presence of spineless eggs across the study area (P = 0.00).

Fig. 3. A typical Schistosoma haematobium egg with the terminal spine.

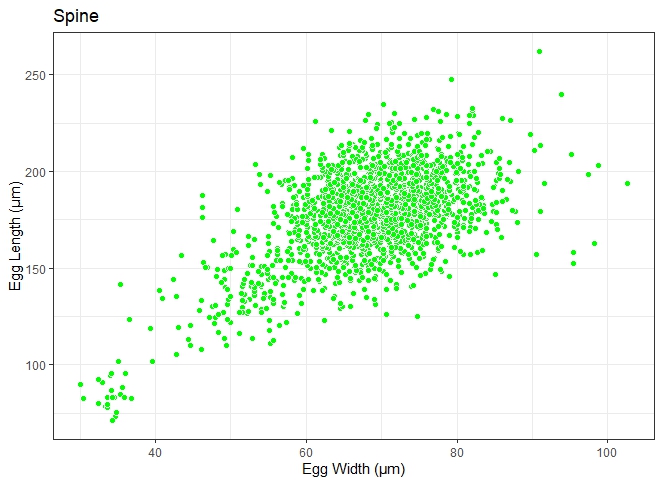
Fig. 4. An atypical spindle-shaped Schistosoma egg.

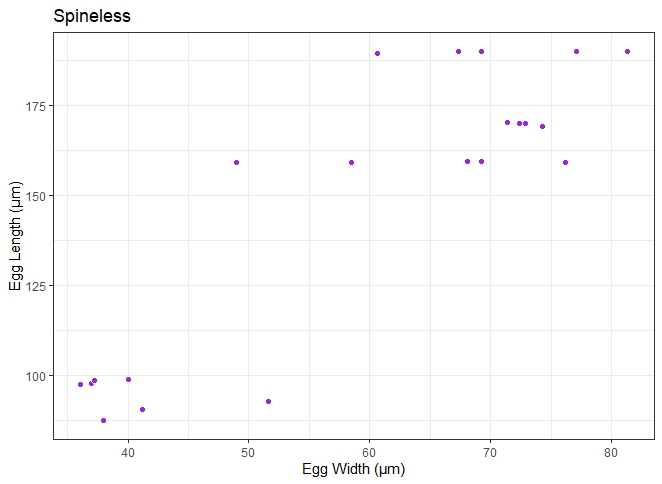
Fig. 5. A spineless Schistosoma egg.
Table 2. The morphotypes of Schistosoma eggs across the study areas.

df, degrees of freedom; *, crosstabulation
Morphometrics of Schistosoma eggs across the study areas
Two major egg morphotypes were identified across the 1984 eggs that were analysed. The first morphotype represented 67.8% of the eggs, with a round-to-oval shape, mean length and width of 166 ± 18 μm and 66.8 ± 9 μm, respectively (fig. 6). Also, the second morphotype represented 32.2% of the eggs and are more elongated, with a mean length and width of 198 ± 18 μm and 71.3 ± 8 μm, respectively (fig. 7). The dimensions of each morphotype were significantly different, both across the communities surveyed, and when grouped (for length: t = −35.374, degrees of freedom (df) = 1982, P = 0.000; for weight: t = −10.431, df = 1982, P = 0.000) The mean length, mean width and length/width ratio of the eggs for both morphotypes are shown in table 3.

Fig. 6. Scatterplot distribution of typical Schistosoma haematobium egg recovered from human urine. Length is within 83–187 μm (Pitchford, Reference Pitchford1965).

Fig. 7. Scatterplot distribution of atypical Schistosoma egg recovered from human urine. Length exceeds 83–187 μm (Pitchford, Reference Pitchford1965).
Table 3. The morphometrics of Schistosoma eggs recovered across the study areas (length of typical Schistosoma haematobium egg: 83–187 μm; Pitchford, Reference Pitchford1965).

Discussion
Human urinary schistosomiasis caused by S. haematobium is one of the most important NTDs in Sub-Saharan Africa, with a wide geographical spread in Africa, the Middle East and Corsica (World Health Organization, 2022). Nigeria remains the most endemic country in Africa, and this present study reports a prevalence of 57.0% (range: 47.1%–63.8%) across the four study communities. This prevalence falls within the World Health Organization hyperendemic thresholds (World Health Organization, 2022), and conforms with earlier reports, highlighting unabated transmission of schistosomiasis across the study area, despite ongoing chemotherapy interventions (Akinwale et al., Reference Akinwale, Ajayi, Akande, Gyang, Adeleke, Adeneye, Adebayo and Dike2010; Ekpo et al., Reference Ekpo, Odeyemi, Sam-Wobo, Onunkwor, Mogaji, Oluwole, Abdussalam and Stothard2017). For over a decade, control efforts have been focused on humans, with little attention given to schistosomiasis in livestock. Schistosoma bovis, a zoonotic species has an extensive geographical distribution throughout Africa, and have been reported in Benin, Togo, Burkina Faso, Niger and Nigeria (Moné et al., Reference Moné, Mouahid and Morand1999; Savassi et al., Reference Savassi, Mouahid, Lasica, Mahaman, Garcia, Courtin, Allienne, Ibikounlé and Moné2020), Countries such as Mali, Senegal, Niger and the Republic of Benin have reported the hybridization of S. haematobium that infects humans and S. bovis that infects cattle (Huyse et al., Reference Huyse, Webster, Geldof, Stothard, Diaw, Polman and Rollinson2009, Reference Huyse, Van Den Broeck, Hellemans, Volckaert and Polman2013; Léger et al., Reference Léger, Garba, Hamidou, Webster, Pennance, Rollinson and Webster2016; Soentjens et al., Reference Soentjens, Cnops, Huyse, Yansouni, De Vos, Bottieau, Clerinx and Van Esbroeck2016; Savassi et al., Reference Savassi, Mouahid, Lasica, Mahaman, Garcia, Courtin, Allienne, Ibikounlé and Moné2020, Reference Savassi, Dobigny, Etougbétché, Avocegan, Quinsou, Gauthier, Ibikounlé, Moné and Mouahid2021). These countries share the Niger river and its tributaries, with Nigeria, where the possibility of migrating snails infected with hybrid schistosomes (S. haematobium x S. bovis) is likely to become established in the Nigerian rivers system. Although snails’ migratory patterns can be influenced by flood, their powers of dispersal are often limited, compared to humans and cattle, who are more mobile species with greater dispersal tendencies, necessary to the spread Schistosoma hybrids. Our study therefore marks as the first attempt to investigate the morphotypes and morphometrics of Schistosoma eggs collected from human samples in Nigeria, as part of an ongoing effort to detect whether hybridization cases already exist in the country.
Currently, there are no studies reporting the spatial co-distribution of S. haematobium and S. bovis. However, S. haematobium has been largely reported among humans in Nigeria (Ekpo et al., Reference Ekpo, Odeyemi, Sam-Wobo, Onunkwor, Mogaji, Oluwole, Abdussalam and Stothard2017; Otuneme et al., Reference Otuneme, Obebe, Sajobi, Akinleye and Faloye2019; Ejike et al., Reference Ejike, Oluwole, Omitola, Bayegun, Shoneye, Akeredolu-Ale, Idowu, Mafiana and Ekpo2021), and there are scanty but emerging reports on S. bovis among livestock (Hambali et al., Reference Hambali, Adamu, Ahmed, Bokko, Mbaya, Tijjani, Biu, Jesse and Ambali2016). Although the eggs of both species are terminally-spined, they differ in several other aspects. For instance, S. haematobium eggs are deposited in urine, and are round-to-oval in shape with a length ranging from 83 to 187 μm (Pitchford, Reference Pitchford1965). In contrast, S. bovis eggs are deposited in stool and are spindle-shaped, consisting of a broad middle portion and drawn-out rod-like ends, one bearing a well differentiated spine, the other evenly rounded (Taylor, Reference Taylor1970; Touassem, Reference Touassem1987). Schistosoma bovis eggs are also larger than those of S. haematobium, with a length ranging from 90 to 220 μm (Pitchford, Reference Pitchford1965), and as high as 300 μm (Taylor, Reference Taylor1970; Touassem, Reference Touassem1987). On the other hand, we found 22 spineless eggs with miracidia, representing approximately 1% of all eggs examined, with the majority of them having the typical S. haematobium shape and size range. It is therefore valuable to further unravel in future studies, the potentials and contributions of these eggs in the S. haematobium and S. bovis hybridization pathway. Nevertheless, about 67% of the egg sizes reported in our study (166⋅84 ± 18.36 μm × 66.82 ± 9.03 μm) corresponds very well with S. haematobium egg sizes (Pitchford, Reference Pitchford1965). In addition, the mean egg length/width ratio of 2.5 recorded for these eggs also corresponds with that reported by Boon et al. (Reference Boon, Fannes, Rombouts, Polman, Volckaert and Huyse2017). However, the remaining one-third of the eggs had an intermediate shape between the typical S. haematobium and S. bovis ova. These eggs are more elongated in shape. These findings conform with the report of Savassi et al. (Reference Savassi, Mouahid, Lasica, Mahaman, Garcia, Courtin, Allienne, Ibikounlé and Moné2020) in the Republic of Benin, and Moné et al. (Reference Moné, Holtfreter and Allienne2015) in France. The sizes of the atypical eggs (198.30 ± 18.82 μm × 71.25 ± 8.44 μm) are significantly longer and bigger than reported for S. haematobium. Also, the 2.8 mean egg length/width ratio recorded is significantly higher than those of S. haematobium and approaches the ratio reported for S. bovis by Boon et al. (Reference Boon, Fannes, Rombouts, Polman, Volckaert and Huyse2017). These dimensions for the intermediate eggs further suggest that these eggs have a shape resembling S. bovis.
The egg shape was taken using digital microscopes and sizes were measured using the IC Measure™ in the laboratory (off-field). Obviously, these morphological methods are time-consuming and hence would be impractical in the context of the high-throughput screening of samples, especially in the field. Besides, microscopic and morphological methods are best suited for rapid detection of eggs and identification of species, mostly for programmatic purposes (Kincaid-Smith et al., Reference Kincaid-Smith, Tracey and de Carvalho2021). However, egg size and shape (length/width ratio) could be highly variable within a single patient, hence impeding species identification (Almeda et al., Reference Almeda, Ascaso, Marçal, Corachan, Southgate and Rollinson1996). As such, morphometric analysis of eggs cannot be solely used as a tool in establishing the presence of hybrid schistosomes (Boon et al., Reference Boon, Fannes, Rombouts, Polman, Volckaert and Huyse2017). It should therefore be noted that this present study did not use molecular methods to verify species status. Hence, some of these measurements could include hybrid rather than pure S. haematobium or S. bovis eggs. It is therefore important to validate our hypothesis of a possible hybridization between S. haematobium x S. bovis with DNA sequencing of eggs from both morphotypes. Molecular analyses are currently being planned; however, the findings reported here provide preliminary evidence on the morphotypes and morphometrics of eggs recovered from human urine in the south-western region of Nigeria, which sheds light on an important but understudied area, hence calling for more research efforts in other parts of the country.
Conclusion
Our results revealed significant variations in sizes of the schistosome eggs recovered, with the atypical shaped eggs appearing more elongated than expected. These eggs might represent individuals with some degree of contribution from S. bovis or possibly other Schistosoma species known to be present in Nigeria. Hence, this observation calls for further molecular studies to establish the genetic information about the miracidia from both atypical and typical eggs. It is also important to establish the presence of bona fide S. bovis infection in cattle and vector snails in the presumptive areas of hybridization.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0022149X22000815
Acknowledgements
We are grateful to the community leaders and residents of the study sites for their continuous participation. Appreciation goes to the Neglected Tropical Diseases Department of the Ogun State Ministry of Health for providing praziquantel used in treating the positive participants. Our profound gratitude goes to the Nigerian Institute of Medical Research, Yaba Lagos, for their support and collaboration in this study.
Financial support
This research was partly funded by Tertiary Education Trust Fund (TETFUND) for Institutional Based Research awarded to U.F. Ekpo under the grant reference number FUNAAB/IBR2019/005.
Conflicts of interest
None
Ethical statement
This study received ethical approval from the Health Ethics Review board of Ogun State Hospital, Ijaiye, with the approval number SHA/RES/VOL.4/154. A pre-survey contact/advocacy meeting was organized to study communities to obtain consents from the community leaders after explaining the objectives of the research to them. Communities willing to participate in the study completed written consent forms. Subsequently, children and adults were also informed about the study through community meetings organized by the consenting representatives. Formal written consents were provided by all participants above 16 years of age. However, for children below 16 years of age, an assent form was completed by each child, in addition to a consent form completed by their parents/caregiver. Only participants with completed consent and assent forms were recruited in the research. The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2008.