



Introduction
Recognizing the first appearance of domestic animals is central to understanding how farming spread across Europe. This is not straightforward: it requires confident identification of domestic taxa, reliable dates, and, perhaps most difficult, providing evidence of actual farming. A series of papers published starting several decades ago raised the possibility of hunted populations of escaped domestic animals confusing the picture of the Neolithization process and potentially playing a starring role (Davidson, Reference Davidson and Clutton-Brock1989; Bogucki, Reference Bogucki and Capana1995, Reference Bogucki1996, Reference Bogucki and Grygiel2008). These hypotheses were rooted in ethnohistoric observations. However, this discussion has largely been ignored in the literature. This is unfortunate because this concept, proposed by Peter Bogucki (Reference Bogucki and Capana1995, Reference Bogucki1996, Reference Bogucki and Grygiel2008) and Iain Davidson (Reference Davidson and Clutton-Brock1989), deserves far more attention. This absence of discussion has also meant that the presence of domestic animals in hunter-gatherer contexts has most often been interpreted as the arrival of farmers and farming, including a degree of husbandry. It is my aim here to revisit this question in more depth to show that this is not necessarily the case because wherever there are farmers, there are feral livestock, and often in large numbers. I also hope to show that hunter-gatherer populations can acquire livestock, but often this has nothing to do with farming and that there may already be evidence of this at the transition to the Neolithic in Europe. This is an extremely challenging aspect of recognizing the arrival of the Neolithic, but it is also an opportunity to understand what the presence of feral animals may have meant. My objective is to chart a path forward by demonstrating that the study of feral animals is integral to understanding transitions to farming and that the meaning of low numbers of morphologically domestic taxa must be more critically assessed.
The eastern seaboard of North America at the time of its colonization by Europeans in the seventeenth and eighteenth centuries provides a good example (see Bogucki, Reference Bogucki and Capana1995) of the opportunities archaeological and ethnohistoric records can offer to illustrate what happens when domestic livestock is introduced to a temperate environment on a continental scale. There have been entire books on the subject (e.g. Anderson, Reference Anderson2004). The perspectives drawn from colonial and indigenous perspectives, both ethnohistorically and archaeologically, provide unparalleled insights into processes that may also have characterized early agricultural Europe. Drawing on this record, and focusing predominantly on British colonization, I review the ethnohistoric literature for records of feral populations and escaped livestock. My review is not exhaustive, but I hope persuasive in demonstrating the widespread occurrence of feral animals in this setting. Where available, I also discuss instances where indigenous peoples obtained domestic animals, and how this is reflected in the zooarchaeological record. Lastly, I use this evidence to lend my voice to arguments that propose that this almost certainly happened in the prehistoric past, and that it must have been a frequent occurrence. A consideration of the implications for our understanding of the Neolithization process concludes this article.
Feralization
Melinda Zeder (Reference Zeder2006) has described the process of domestication as proceeding along a spectrum of coadaptation which can result in the domestication of an animal. However, an opposite process also occurs, the process of feralization. Feral animals have been defined as ‘free-living organisms or populations that are primarily descended from domesticated ancestors’ (Gering et al., Reference Gering, Incorvaia, Henriksen, Conner, Getty and Wright2019: 1138). Domestic animals that are unnaturally introduced to a new habitat and have escaped are termed ‘feral exotic livestock’ (see McKnight, Reference McKnight1964). Such animals are exceptionally common; there are numerous modern examples of feral exotic livestock populations in North America (McKnight, Reference McKnight1964), Europe (Kugler & Broxham, Reference Kugler and Broxham2014), Australia (McKnight, Reference McKnight1976), Asia (Hoffman, Reference Hoffman2010), South America (Rodriguez & Martinez, Reference Rodriguez and Martinez1992), Africa (Muller & Bourne, Reference Muller and Bourne2018), and even sub-Antarctic islands (Micol & Jouventin, Reference Micol and Jouventin1995). Feralization can result from various processes, including abandonment, free-range husbandry, and deliberate loss, but the most common origin is accidental escapes (McKnight, Reference McKnight1964: 60). This may have been caused by a number of factors, including poorly maintained or damaged enclosures, human error, or simply clever animals (McKnight, Reference McKnight1964; Takahashi & Tisdell, Reference Takahashi and Tisdell1992; Fraser et al., Reference Fraser, Cone and Whitford2000). Therefore, everywhere domestic animals have been introduced, there have been individuals or populations that have become feral, and this happens regardless of the husbandry method employed.
The recognition that feral exotic livestock are a universal phenomenon creates a conundrum for archaeologists. This is particularly true for those studying the transition to the Neolithic. More specifically here, it is a problem for the study of the arrival of animal husbandry and how we may distinguish between feral and domestic animals in the archaeological record. For my purposes, it is necessary to broaden the definition given above by Gering et al. (Reference Gering, Incorvaia, Henriksen, Conner, Getty and Wright2019) to include animals which have escaped and are not under the day-to-day control of farmers. They may not necessarily form breeding populations but survive without human intervention. Livestock identified as domestic in archaeological contexts could in fact be feral or may have interacted with feral animals or their lineages.
Numerous taxa can go feral (McKnight, Reference McKnight1964), but the most important are those initially domesticated in the Near East, which spread to Europe and have a primary function in agricultural production (Manning et al., Reference Manning, Downey, Colledge, Conolly, Stopp and Dobney2013a; McClure, Reference McClure2015; Vigne, Reference Vigne2015). These include the domestic cow (Bos taurus), pig (Sus scrofa), sheep (Ovis aries), and goat (Capra hircus). To these I add horses (Equus caballus) because, although their domestication postdates the European Neolithic (Vigne, Reference Vigne2015), the taxon is relevant to the North American examples given below. Other domestic animals can go feral, including domestic dogs (Canis familiaris) and cats (Felis silvestris), but neither taxon is generally considered to play a central role in farming.
Feral animals retain the morphological traits of the domestic form, especially in pigs, although feralization may change the size of individuals (Neaux et al., Reference Neaux, Sansalone, Lecompte, Haruda, Schafberg and Cucchi2020). Depending on the setting, there will sometimes be rapid selection and adaptation to new environments. The feral cattle on the isolated Amsterdam Island in the southern Indian Ocean, for example, have lost about a quarter of their overall body size in around a century on the island (Rozzi & Lomolino, Reference Rozzi and Lomolino2017). A similar effect has been noted archaeologically: there were probably some morphometric changes in domestic cattle that accompanied agricultural colonization in eastern North America (Arbuckle & Bowen, Reference Arbuckle, Bowen, Mondini, Munoz and Wickler2004); most are likely to have been caused by adaptation to new environments. Therefore, recognizing feral animals on morphological and/or morphometric grounds will not be straightforward, because the new environment may affect the animals’ size, whether they are under human control or feral, and feral animals may retain domestic features. The upshot is that recently feralized animals will probably continue to look ‘domestic’ in the archaeological record, except in instances where long-term interbreeding with wild populations results in a ‘wild’ phenotype despite retaining a partial domestic genotype (e.g. Rowley-Conwy & Zeder, Reference Rowley-Conwy and Zeder2014).
Feralization in North America
The earliest European livestock was introduced to mainland North and Central America at the same time as, or shortly after, the arrival of Europeans (Bowling, Reference Bowling1942). What happened next is what matters to us: they promptly escaped (Stewart, Reference Stewart1991) or were allowed to roam free. As part of a broader pattern in eastern North America, in the Chesapeake region of Virginia (Figure 1), British colonists discovered that an abundance of land and labour being at a premium necessitated free-range husbandry methods: animals were not kept penned but were allowed to roam freely in the woods (Anderson, Reference Anderson2002, Reference Anderson2004). One result was that feral breeding populations were quickly established. A Virginia census of ad 1620, scarcely more than a decade after the arrival of the British, even struggled to quantify the large numbers of feral pigs in the forest (Anderson, Reference Anderson2002: 400). The feral swine did not escape the notice of the indigenous Native American population, and in Virginia Captain John Smith noted that ‘…wilde hogs, which were infinite, are destroyed and eaten…’ by Native Americans (Smith, Reference Smith1910 [1627]: 885). As time passed, the presence of feral animals in the woods became common across the colonies (Gray & Thompson, Reference Gray and Thompson1933: 140) and numbers increased. To wit, by the mid-seventeenth century in the Virginia Colony, there were more wild (feral) cattle than domestic cows (Bowling, Reference Bowling1942). In South Carolina, John Smyth (d. ad 1682), a cattle farmer, reported that in addition to his fifty-odd cattle, there were around thirty running wild in the woods (cited in Otto, Reference Otto1987: 19). In some cases, the animals spread in advance of stockkeeping: for example, when British settlers arrived in Georgia, there were already large numbers of feral cattle there, derived at least in part from Spanish cattle introduced some time earlier, and they had spread north from what is today Florida (Stewart, Reference Stewart1991; Bogucki, Reference Bogucki and Capana1995).

Figure 1. Map of eastern North America with regions and sites mentioned in the text. Locations are approximate. Basemap from the GADM database (www.gadm.org), version 3.4, April 2018.
Ultimately, the feral exotic livestock became so ubiquitous that they were validated by legislation and in sport. So bad was the problem that the General Assembly of Maryland decreed in ad 1661, ‘And it is further Enacted that it shall be lawfull after the end of february vntill the first of december for any Inhabitant of this Province to kill any wilde vnmarked Cattle resorteing in Company with their tame Cattle’ (General Assembly of Maryland, 1661: 418–19). Similar legislation was passed elsewhere in the colonies (Gray & Thompson, Reference Gray and Thompson1933). Additionally, so widespread were feral animals in the woods that, by the earlier eighteenth century, hunting of horses was carried out for sport in Virginia, there being so many wild horses in the uplands (Beverley, Reference Beverley1722; Gray & Thompson, Reference Gray and Thompson1933). Thus, upon introduction to the Americas, exotic livestock were soon living and breeding as feral populations; this was a widespread phenomenon, and one extensively documented in the historical and ethnohistoric literature (Bogucki, Reference Bogucki and Capana1995; Gibson, Reference Gibson2016).
The North American Archaeological Record
The questions of whether exotic domestic livestock were acquired by Native Americans, how and why this happened, and how this is represented in the zooarchaeological record, are all relevant for comparison with the process of Neolithization in northern and western Europe. However, post-colonization Native American subsistence practices are an under-studied aspect of historical archaeology (Landon, Reference Landon2005). Nonetheless, in order to understand whether domestic animals made it into Native American possession, the study of post-colonization Native American assemblages is necessary.
It must first be acknowledged that the Native American residents of North America represent a wide variety of cultures, each with their own attitudes towards subsistence, the local environment, exotic animals, and almost certainly several other unknown factors. Furthermore, they were aware of agriculture, actively practised farming (Doolittle, Reference Doolittle1992; Scarry & Scarry, Reference Scarry and Scarry2005), possessed domestic dogs (Derr, Reference Derr2004), and perhaps managed turkeys (Peres & Ledford, Reference Peres and Ledford2016). The implications of this knowledge on potential receptivity of indigenous groups to livestock husbandry is, however, largely unclear because the first encounters with exotic livestock were characterized by a mix of fascination, surprise, and sometimes fear, and were followed by attempts to fit the exotic livestock into existing worldviews regarding hunted animals (Anderson, Reference Anderson2002, Reference Anderson2004; for a similar example from Australia, see May et al., Reference May, Wright, Sanz, Goldhahn and Maralngurra2020). Livestock husbandry could at times also be incompatible with existing land-use preferences (Pavao-Zuckerman, Reference Pavao-Zuckerman2007). Moreover, the force of colonial power dynamics meant that existing Native American political structures became intertwined with those of the colonists, resulting in diverse situations in which Native Americans were variously encouraged to acquire exotic taxa, or were prevented from doing so, or desired or avoided them (Salisbury, Reference Salisbury1996; Anderson, Reference Anderson2002; Peres & Ledford, Reference Peres and Ledford2016). It is therefore probably impossible to speak in anything but the most general terms about the role exotic domestic animals played in their lives. We must also remember that the presence of domesticated animals in Native American faunal assemblages does not even indicate that there were necessarily stockkeepers nearby. In fact, it seems that exotic taxa were present earlier than sustained or even indirect contact with incoming European populations (Pavao-Zuckerman & Reitz, Reference Pavao-Zuckerman and Reitz2006).
Nonetheless, post-colonization, there is generally a low contribution of exotics in Native American faunal assemblages (Figure 2). Faunal assemblages from Colonial period Mohawk sites in central New York State contained European domesticates alongside wild taxa, and their presence is documented by the end of the seventeenth century (Kuhn & Funk, Reference Kuhn and Funk2000). Similarly, there were low numbers of domestic animals, mostly pigs, in the post-colonization (c. ad 1600–1819) Cherokee settlement Toqua in Tennessee (Bogan, Reference Bogan1980; Pavao-Zuckerman, Reference Pavao-Zuckerman2000). At the Seneca Iroquois Townley-Read site (c. ad 1715–1754) in New York State the faunal assemblage is dominated by wild taxa with just a few specimens of cattle and pigs (Watson & Thomas, Reference Watson and Thomas2013). Perhaps the best perspective comes from the Creek town of Fusihatchee, in what is today Alabama, where the faunal assemblage spans both pre- and post-colonization periods. Importantly, the protohistoric components show cultural conservatism with regard to the incorporation of European domestic animals; nevertheless domestic animals of European origin are present in low numbers in the seventeenth century (Pavao-Zuckerman et al., Reference Pavao-Zuckerman, Weinand and Reitz1999). The breakdown of wild versus exotic livestock from several of the sites noted above is shown in Figure 2.

Figure 2. Post-colonization Native American faunal assemblages c. 1620–1820, by per cent Number of Identified Specimens (NISP). Data based on Bogan, Reference Bogan1980 (c. 1600–1819 data only), Pavao-Zuckerman et al., Reference Pavao-Zuckerman, Weinand and Reitz1999, Pavao-Zuckerman, Reference Pavao-Zuckerman2000, and Watson & Thomas, Reference Watson and Thomas2013. Birds, fish, bats, amphibians, invertebrates, reptiles, small rodents (rats, voles, shrews, chipmunks, mice), dogs, and wolves are omitted.
How do the exotic livestock make their way into these assemblages? Their presence in protohistoric Fusihatchee has been interpreted as owing to trade, raiding, or hunting of feral animals (Pavao-Zuckerman, Reference Pavao-Zuckerman2007). Similarly, some scholars argue that the presence of pig remains in the Seneca assemblages ‘…more likely represented the introduction of another source of wild meat into the ecosystem…’ (Jordan, Reference Jordan2008: 295). It could also be because of conflict. In New Netherlands (New York), it was reported that ‘…the cattle usually roamed through the woods without a herdsman, they frequently came into the corn of the Indians which was unfenced…this led to frequent complains on their part and finally to revenge on the cattle without sparing even the horses…’ (Anonymous, Reference Jameson1649 cited in Jameson, 1909: 273).
Exotic livestock is just as often absent in post-colonization Native American assemblages. Contact-period Sara sites in North Carolina, for example, show no incorporation of any exotic livestock, with subsistence largely unchanged from pre-colonization periods, a pattern of continuity which is similar across the south-eastern part of North America (Pavao-Zuckerman, Reference Pavao-Zuckerman2000; Holm, Reference Holm, Eastman, Rodning and Boudreaux2002). Overall, this record illustrates that there is no widespread adoption or husbandry of exotic livestock immediately after contact, but that it is relatively common for small numbers of their bones to be present in archaeological assemblages.
The Neolithization of Northern and Western Europe
Numerous problems surround the identification of the earliest exotic livestock in Stone Age northern Europe. The most important are those associated with taxonomic identification, of which there is a variety of methods of varying accuracy (e.g. Scheu et al., Reference Scheu, Hartz, Schmölcke, Tresset, Burger and Bollongino2008; Rowley-Conwy et al., Reference Rowley-Conwy, Albarella and Dobney2012; Evin et al., Reference Evin, Cucchi, Cardini, Vidarsdottir, Larson and Dobney2013; Owen et al., Reference Owen, Dobney, Evin, Cucchi, Larson and Vidarsdottir2014). Even if the presence of exotic livestock can be confirmed, it is clear that the mere presence of such animals does not mean the Neolithic has started. Given the abundant evidence of the modern and historical occurrence of feral exotic livestock and their low-level presence in indigenous faunal assemblages, feral, stray, or escaped animals are likely to have been a feature of the Neolithic in northern and western Europe (Bogucki, Reference Bogucki and Capana1995). Their presence in low numbers in faunal assemblages therefore need not suggest active husbandry at a given site. Such an understanding helps us understand interpretive disagreements and conundrums regarding fast versus slow introductions of farming practices in northern and western Europe (see Gron & Sørensen, Reference Gron and Sørensen2018; Crombé et al., Reference Crombé, Aluwé, Boudin, Snoeck, Messiaen and Teetaert2020). We may need to rethink our understanding of how we date the arrival of agriculture, because the North American example demonstrates the ‘first’ or ‘earliest’ domestic animals in a secondary context may in fact have preceded actual animal husbandry, or even actual stockkeepers on the landscape.
The possibility that feral animals were hunted is sometimes mentioned, but often dismissed in the archaeological literature, usually in order to claim Mesolithic management of taxa or to explain otherwise anachronistic specimens. In western France, for example, there are several sites of Mesolithic character which have yielded low numbers of domestic animals, suggested as having been obtained ‘…through exchange with agriculturalists, the theft of animals, or the hunting of strays’ (Arias, Reference Arias1999: 428). Contentious claims (see Rowley-Conwy & Zeder, Reference Rowley-Conwy and Zeder2014) that Ertebølle foragers kept domestic pigs on the basis of aDNA evidence (Krause-Kyora et al., Reference Krause-Kyora, Makarewicz, Evin, Flink, Dobney and Larson2013) can be explained if we consider that feral pigs are likely to have interbred with wild boar, something aDNA analysis has confirmed was common throughout Europe (Frantz et al., Reference Frantz, Haile, Lin and Scheu2019).
Recently, domestic animal husbandry has been suggested in a hunter-gatherer context at the site of Bazel in north-western Belgium (location on Figure 3). This claim has proven contentious among zooarchaeologists (e.g. Brusgaard et al., Reference Brusgaard, Çakirlar, Dee, Dreshaj, Erven, Peeters and Raemaekers2022). Despite unclear stratigraphy, direct radiocarbon dates of both domestic cereals and exotic livestock, some archaeologists have suggested the presence of small-scale animal husbandry early in the occupational sequence (Meylemans et al., Reference Meylemans, Bastiaens, Boudin, Deforce, Ervynck and Perdaen2018; Crombé et al., Reference Crombé, Aluwé, Boudin, Snoeck, Messiaen and Teetaert2020). In these early phases, the cereals have been interpreted as traded to the hunter-gatherers, as there is no evidence for local cultivation in the pollen data (Meylemans et al., Reference Meylemans, Bastiaens, Boudin, Deforce, Ervynck and Perdaen2018: 5). The possibility of the identified animals being feral was dismissed on the grounds that there were multiple domestic taxa present in the assemblage (Crombé et al., Reference Crombé, Aluwé, Boudin, Snoeck, Messiaen and Teetaert2020). The proportion of wild versus domestic taxa is, however, unclear and, as previously explained, multiple exotic livestock can go feral in one place at one time. Furthermore, the Bazel cattle have bone collagen δ13C isotope values (almost all lower than c. -22.6‰ δ13C, and sometimes much more so) consistent with a forest habitat (Gron & Rowley-Conwy, Reference Gron and Rowley-Conwy2017; Gron et al., Reference Gron, Rowley-Conwy, Fernandez-Dominguez, Gröcke, Montgomery, Nowell and Patterson2018). This is unlike other instances in northern and western Europe, where it is clear that domestic cattle were controlled and raised in open environments (Figure 4). The Bazel animals could indeed have been feral, and similarity between the isotope values of many of the cattle, red deer, and aurochs at Bazel may be interpreted as evidence that the cows were feral. If this is the case, this substantially alters the interpretation of the earliest phases at the site, and therefore the local process of Neolithization. Instead of representing small-scale husbandry, the earliest phases could represent a time when local hunter-gatherers were trading with adjacent farmers for cereals, and occasionally hunted feral exotic livestock in the surroundings.
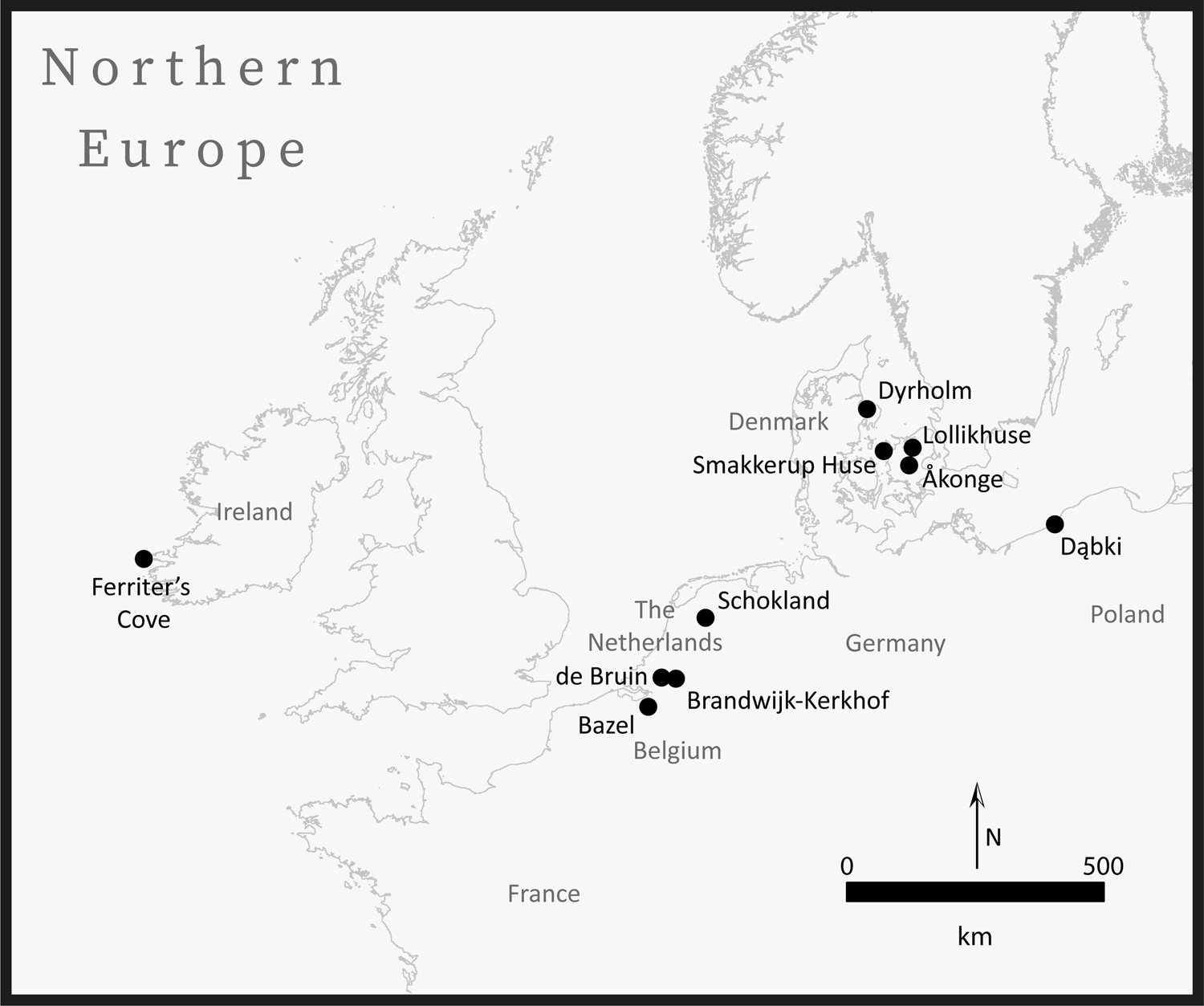
Figure 3. Map of northern Europe with later prehistoric site locations (approximate). Basemap from the GADM database (www.gadm.org), version 3.4, April 2018.

Figure 4. Carbon and nitrogen stable isotope evidence for browsing environments of herbivores in later prehistoric western and northern Europe. Data based on Meylemans et al., Reference Meylemans, Perdaen, Sergant, Bastiaens, Crombé and Debruyne2016, Gron & Rowley-Conwy, Reference Gron and Rowley-Conwy2017, Gron et al., Reference Gron, Rowley-Conwy, Fernandez-Dominguez, Gröcke, Montgomery, Nowell and Patterson2018, Crombé et al., Reference Crombé, Aluwé, Boudin, Snoeck, Messiaen and Teetaert2020, and Gron, Reference Gron, Gron, Sørensen and Rowley-Conwy2020. Uncertain or mixed taxonomic identifications of Bos sp. are omitted. The dotted line represents the approximate boundary between grazing environments.
Elsewhere, other early domesticated animals may also have been feral, especially when their incidence is low relative to wild taxa. This includes early cattle, pigs, and caprines at de Bruin and Brandwijk-Kerkhof (both in the Netherlands) (Çakırlar et al., Reference Çakırlar, Breider, Koolstra, Cohen, Raemaekers, Gron, Sørensen and Rowley-Conwy2020), cattle and pigs at Schokland (also in the Netherlands) (Gehasse, Reference Gehasse1995), possible cattle and pigs at Dąbki (Poland) (Schmölcke & Nikulina, Reference Schmölcke, Nikulina, Kabaciński, Hartz, Raemaekers and Terberger2015), as well as cattle at Dyrholm, Smakkerup Huse, and Åkonge (Denmark) (Rowley-Conwy, Reference Rowley-Conwy, Colledge, Conolly, Dobney, Manning and Shennan2013) (locations in Figure 3). In all these cases, the few domesticates, even if identifications and dates are proven secure, could be explained as feral, escaped, or otherwise obtained.
The only exceptions, i.e. where feral animals cannot be implicated, are the rare instances of exotic livestock arriving, by whatever means, to islands too distant to reach by swimming or travelling over ice in the winter. Two notable examples include the possible early cow from Lollikhuse in Zealand (Denmark) pre-dating the start of the Neolithic by nearly 800 years (Sørensen, Reference Sørensen and Bujnte2005) and the early cattle from Ferriter's Cove, Ireland (Woodman et al., Reference Woodman, Andersen and Finlay1999) (location in Figure 3). In these cases, some other process must be at play; for Ferriter's Cove, the prevailing interpretation is that the cattle were exotic livestock from a failed Neolithization episode that had been hunted by indigenous foragers (Sheridan, Reference Sheridan, Finlayson and Warren2010). Be that as it may, low incidences of domesticates certainly do not demonstrate local animal husbandry at these sites.
Conversely, why are there wild animals in clearly agrarian contexts? Some earlier Neolithic faunal assemblages in several locations across Europe, which are clearly agrarian in character, are notable for the substantial contribution of wild taxa (Manning et al., Reference Manning, Downey, Colledge, Conolly, Stopp and Dobney2013a, Reference Manning, Stopp, Colledge, Downey, Conolly, Dobney, Colledge, Conolly, Dobney, Manning and Shennan2013b). It is likely that this indicates the ‘persistence of high numbers of wild taxa during the early Neolithic’ (Manning et al., Reference Manning, Downey, Colledge, Conolly, Stopp and Dobney2013a: 1051) in the environment, a process also identified among incoming Europeans in the Chesapeake frontier in seventeenth-century North America (Miller, Reference Miller1984). In the Chesapeake case, the earliest incoming stockkeepers incorporated abundant wild taxa in the earliest assemblages, a combination that gradually waned as the numbers of settlers increased, affecting the balance of the environment and of the native fauna (Figure 5). The idea of domestic animals affecting, even shaping, environments in Neolithic Europe is not new, since it has been suggested that animal husbandry is a form of Neolithic niche construction (McClure, Reference McClure2015). This certainly occurred in the colonial Chesapeake region (Curtain et al., Reference Curtain, Brush and Fisher2001; Hall, Reference Hall2018), with animals actively used to shape the environment.

Figure 5. Early Chesapeake settlement assemblages c. ad 1620–1720 (left) and European LBK assemblages (right), by per cent NISP. Sites are arranged for each period in chronological order with the earliest on the left, with some overlap in dates. Data based on Barber, Reference Barber1978 (cited in Miller, Reference Miller1984: 395, tab. 38), Bowen, Reference Bowen and Nieman1979 (cited in Miller, Reference Miller1984: 409, tab. 50), Miller, Reference Miller1984, and Manning et al., Reference Manning, Stopp, Colledge, Downey, Conolly, Dobney, Colledge, Conolly, Dobney, Manning and Shennan2013b. Birds, fish, domestic cats, amphibians, crustaceans, rats, dogs, and wolves are omitted. The numbers above columns refer to the sample size.
Discussion
The overwhelming ethnographic and historical evidence for feral animal populations from the colonial contact period of eastern North America, and the nearly ubiquitous presence of feral animals wherever introduced worldwide, suggest that they were almost certainly present in previously unfarmed landscapes during the European later Stone Age. This raises substantial doubts as to their meaning when encountered in low numbers in archaeological assemblages. In these cases, it cannot be assumed that they were part of a farming economy, or even that there were farmers nearby. The incorporation of substantial amounts of wild fauna in the earliest farming contexts similarly does not necessarily represent a ‘transition’ to farming lifeways; it can just as plausibly be evidence for incoming farmers exploiting the abundant wild resources of a previously unfarmed landscape.
If we accept that feral animals were present, how do we then identify animal husbandry in the archaeological record? The answer certainly lies in degree. The more contentious claims for small-scale husbandry nearly all rely on evidence that can just as plausibly be explained by the presence of feral animals nearby. But when is the evidence of active husbandry reliable? Clearly, single finds of exotic livestock in Mesolithic contexts (e.g. Sørensen, Reference Sørensen and Bujnte2005) do not mean farming has started. Similarly, nobody would argue that a large quantity of cattle bones at a given site, representing hundreds of individuals, could derive from the hunting of feral animals by hunter-gatherers. It is not my aim here to set out specific interpretive criteria for either because everything will depend on context. A good starting point, however, would be to ask a simple question: can the evidence be reasonably interpreted as deriving from a farming system including crop cultivation and animal husbandry? In other words, are there multiple lines of evidence to suggest active, ongoing farming? If not, alternative explanations, including the involvement of feral animals, need be considered.
We are not without the appropriate methods to develop criteria in the future, especially with regard to the identification of feral animals. Some of the considerable zooarchaeological toolkit that could be applied includes established basic and derived proportional quantitative statistics (e.g. Number of Identified Specimens (NISP), Minimum Number of Individuals (MNI), biomass, and Percent Minimal Animal Units (%MAU)) (see Lyman, Reference Lyman1994). Using these routine data, feral animals obtained by indigenous foragers could be expected to be represented as a relatively small proportion of a faunal economy dominated by wild taxa, and if the animal is larger (i.e. cattle), there should be evidence of selective body-part representation indicative of transport after butchery. Other methods could also be brought to bear, such as stable and radiogenic isotope analysis, aDNA determinations, and other applications from the archaeological sciences. In line with the Bazel example, a population of feral livestock in northern and western Europe can be expected to have isotopically determined diets similar or overlapping with those of contemporary wild herbivore populations. Inbreeding between wild and domestic forms should also be expected, and indeed there is already aDNA evidence to suggest that this occurred (e.g. Rowley-Conwy & Zeder, Reference Rowley-Conwy and Zeder2014). Future research that recognizes the presence of feral animals will enable data generation which will in turn allow their identification but also the confirmation of active animal husbandry at the start of the Neolithic.
Lastly, the feral animal question underscores the value of consulting the ethnographic record before proposing low-level livestock husbandry by a population transitioning to agriculture. Basic but fundamental insights can be gained. How else would we know that by and large the historic-period Native American Creeks of the south-eastern region of eastern North America had no interest in keeping European-introduced cattle simply because they were lactose intolerant (Pavao-Zuckerman, Reference Pavao-Zuckerman2007: 26-27)? Facts like these prompt us to question whether low-level husbandry would even have been a viable economic option for European hunter-gatherers confronted with Neolithization.
Conclusion
It has been my purpose in this contribution to argue for the near certain presence of feral exotic livestock on the European continent in Neolithic times. These animals may, indeed probably were, likely to have been encountered by the last foragers before they ever saw a farmer. The ethnographic record of North America and elsewhere attests to indigenous people obtaining exotic livestock, but this often has little to do with indigenous farming practices. Similarly, incoming Early Neolithic farmers should be expected to be incorporating wild resources as an opportunistic use of abundant resources, but not as a substitute for farming as the basis of the economy. Therefore, when making claims for the existence of the ‘earliest’ domestic animals, or the presence of domestic taxa within hunter-gatherer contexts, or identifying early animal husbandry, a simple question needs to be asked: could these be feral animals? The answer more often than not is probably yes.
Acknowledgements
The Leverhulme Trust (Grant RPG-2019-253) provided support during the preparation of this manuscript. I would also like to thank Valerie Hall, Peter Rowley-Conwy, Jake Pfaffenroth, and Harry Robson.






 Open access
Open access