

Introduction
The three species of manatees (Trichechus spp.) are aquatic mammals of the Order Sirenia. There are conservation concerns for all three species because of collisions with boats, entanglement in fishing nets, hunting and habitat degradation, and all are categorized as Vulnerable on the IUCN Red List (IUCN, 2008).
Similar to the Florida manatee Trichechus manatus latirostris, the Antillean manatee Trichechus manatus manatus lives in salty, brackish and freshwater systems, sometimes alternating between the three environments (Lefebvre et al., Reference Lefebvre, Marmontel, Reid, Rathbun, Domning, Woods and Sergile2001). Although it is known that some Antillean manatee populations live exclusively in the freshwater of the Orinoco River (O'Shea et al., Reference O'Shea, Correa-Viana, Ludlow and Robinson1988; Correa-Viana et al., Reference Correa-Viana, O'Shea, Ludlow and Robinson1990), most studies of the biology and ecology of the species have examined populations that live in coastal waters and estuaries (O'Shea & Ackerman, Reference O'Shea, Ackerman, O'Shea, Ackerman and Percival1995; Flamm et al., Reference Flamm, Ward and Weigle2001). We therefore studied an Antillean manatee population located > 1,100 km from the Caribbean coast, in the Orinoco River of Colombia, from February 2001 to April 2005. The study involved interviewing local inhabitants and making direct observations and records of manatee feeding signs. In particular we aimed to identify areas used by the manatee in different seasons, describe any local migrations, and develop a simple model of habitat use in relation to mortality. Our overall aim was to provide information for the elaboration of management strategies for the species.
Study area
The study was carried out in the Orinoquía region, department of Vichada, Colombia, on the border with Venezuela, in the Reserva de Biósfera El Tuparro. The Orinoco is a clear-water, alkaline, tropical floodplain river system, with low levels of nutrients (Colonello, Reference Colonello1990). Temperature is high throughout the year (mean 28.1°C, range 18.0–42.8°C). There is a dry season from December to March, and the rainy period starts in April (Vasquez & Wilbert, Reference Vasquez, Wilbert, Calow and Petts1992). Because of this variation in annual precipitation there is a marked change in the level of the Orinoco River and its tributaries (Vasquez & Wilbert, Reference Vasquez, Wilbert, Calow and Petts1992), resulting in a unimodal seasonal inundation that usually lasts for 4 months (Hamilton & Lewis, 1990). The low water period is usually from January to April, with the lowest water level in March. The water rises in May–June, and the high water period is in July–October, with a peak in August. The amplitude between the low and high water periods is c. 13 m (Vasquez & Wilbert, Reference Vasquez, Wilbert, Calow and Petts1992). The river level falls in November–December. The flooded areas of the Orinoco are similar to other hydrographic systems, such as the Amazon, in amplitude, duration and ecological characteristics (Junk et al., Reference Junk, Bayley, Sparks and Dodge1989; Hamilton & Lewis, 1990). Changes in the Orinoco River level in 2001–2004 are illustrated in Fig. 1. The study covered c. 40 km of the Orinoco River, (Fig. 2) from El Burro (6°12′ N, 67°26′ W) to Bachaco (5°55′ N, 67°31′ W).
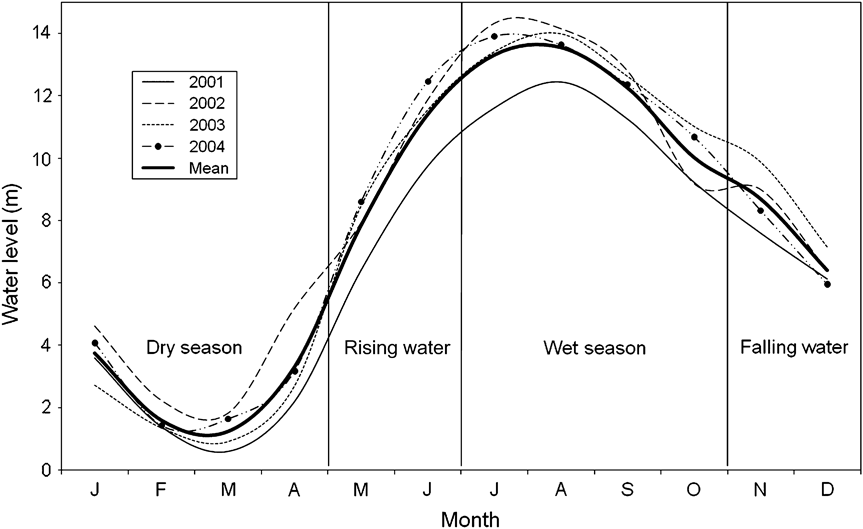
Fig. 1 Hydrological dynamics of the Orinoco River during 2001–2004 (data for 2005 were not available). Data are from Font Instituto de Hidrología, Metereología y Estudios Hidrográficos de Colombia.

Fig. 2 The study area in the Orinoco River of Colombia. Rectangles represent areas where manatees were located (by sightings, feeding signs or interviewee reports of dead or injured manatees, see text for further details) during the (a) low water periods, (b) rising water periods, (c) high water periods and (d) falling water periods of 2001–2005.
Methods
Observations for manatees were carried out in 2001 (low water), 2002 (low and rising water), 2003 (low, rising and high water), 2004 (high and falling water) and 2005 (low water). Eight observation areas were chosen based on information about manatee presence provided by local inhabitants (Punta Lajas, Puerto Sucre, San José, Ventanas, Isla Santa Helena, La Angela, Isla Charal and Tronconal). Water depth was measured at three points in each area. Forest canopy and extreme seasonal water level changes make aerial surveys ineffective, and therefore observations were made during 6.30–17.00 for a minimum of 2 hours on each occasion. In areas such as the Orinoco River a combination of boat and land-based surveys are the only means of monitoring manatees. Four sighting methods were used: from high points on the river margins with a wide view of the area, from a small boat drifting with its engine off (average speed 4.0 km h-1), from an anchored boat, and from the river margins whilst walking. The total number of hours of observation at each site in each year are given in Table 1. Observations were suspended if either weather or waves would have interfered with data accuracy. Sightings were made when the manatees came up to breathe, when they expose their snout, back and sometimes their tail in a conspicuous way. Number of sightings per hour and maximum number of simultaneous sightings (manatees usually breathe simultaneously when in a group, and the maximum number of simultaneous sightings is the maximum number of manatees counted at the surface at one time) were used as relative indices of occurrence. Observations were also made of typical manatee feeding signs on leaves and stalks, which are easily distinguishable by experienced observers because there is no other aquatic herbivore that makes the same marks.
Table 1 Effort, in hours, of observations of manatees in eight areas of the Orinoco River (Fig. 2) during 2001–2005. For each site only years with observations are indicated.

In 2001 (rising water period), 2002 (rising water period), 2003 (rising and high water periods) and 2004 (high and decreasing water periods) we searched for manatee feeding areas and made weekly observations for characteristic feeding signs, using transects, from a small boat with an engine travelling at an average speed of 8 km h-1. We made eight, eight, 16 and 17 transects at each of the eight sites in 2001, 2002, 2003 and 2004, respectively, except at the Punta Lajas site in 2004 where we made nine transects.
During 2001–2005 we carried out a total of 82 interviews in the cities of Puerto Carreño (Colombia) and Puerto Ayacucho (Venezuela) and in 28 fishing communities along the Orinoco and Meta River margins. Interviewees were men, without age restriction, who had some kind of contact with the species. For every recorded occurrence of a manatee killed, wounded, caught in a net or stranded, size class was determined according to weight and/or length estimated by the interviewee, as a calf (< 200 kg and/or < 175 cm), subadult (200–300 kg and/or 176–274 cm) or adult (> 300 kg and/or > 275 cm). These classes are based on size-specific reproduction data estimated for the species (Marmontel, Reference Marmontel1993). Data were rejected if information about the period or location were not provided, if the information was not from the Orinoco River, or if the report pre-dated 1980 (to reduce the risk of errors arising from the passage of time). Reports from different sources were compared to avoid repetitions. If two or more reports had data that coincided, they were considered as the same event. When necessary, interviewees were contacted again to verify information.
Results
In 1,002.5 hours of observation we sighted 870 manatees, an average of 0.87 per hour (Table 2) at Punta Lajas, Puerto Sucre, San José, Ventanas, Isla Santa Helena and Tronconal (Fig. 2). The maximum number of individuals observed simultaneously was 17, in the rising water period of 2002 at Tronconal. Even though observations were made throughout the various hydroclimatic periods, manatees were only observed during the low and rising water periods (January–July) in water depths of 2.0–21.2 m. After 776 hours searching for feeding signs, 69 manatee feeding areas were located, at San José and La Angela, and near the islands of Chimborazo, El Índio, Charal and Playa Caimanes (Table 3). All of the feeding areas were located during the rising and high water periods.
Table 2 Total observation effort, number of sightings per year and total, number of sightings per hour and maximum number of simultaneous sightings (see text for details) of manatees in the study area (Fig. 2) between 2001 and 2005, during periods of low, rising, high and falling water.

Table 3 Effort (in hours) spent searching for manatee feeding areas in 2001–2004 in the study area (Fig. 2), number of feeding areas located, and minimum and maximum water depth at feeding locations.

From the interviews a total of 144 dead and wounded manatees were reported. Following validation the number of cases reduced to 92 during 1980–2004 (Table 4). The most common causes of mortality or wounding were incidental drowning in fishing nets (43.3%) and hunting with harpoons (38.9%). Cases of beach stranding (5.6%), floating or stranded carcasses (7.8%), bullet wounds (2.2%) and collision with boats with engines (2.2%) were also reported. Of the 90 cases, 37 dead or injured manatees were in the study area.
Table 4 Number of cases of dead or injured manatees in the Orinoco River during 1980–2004 based on information obtained in interviews with local people.

* Marmontel (Reference Marmontel1993), see text for further details
In total, from sightings, feeding areas and reports of mortality, we located manatees in 14 locations in the study area (Fig. 2, Table 5). During the low water period manatees were observed in a continuous area from Punta Lajas to the tip of Isla Santa Helena. The places where the individuals were observed during the dry period are known locally as buceaderos, zones that still have a high water level during this time. When the river started rising sightings were made and feeding areas located from San José to Isla El Índio. La Angela, Isla Charal and Tronconal were important feeding sites during this period. In the high water period evidence of feeding was found in La Angela and around the islands of Chimborazo, Playa Caimanes and El Índio. In the decreasing water period we did not locate any feeding areas or manatees but two cases of dead animals were reported at Punta Lajas and Bachaco.
Table 5 Occurrence of manatees at 14 locations in the study area (Fig. 2) during 2001–2005.

* F, feeding areas; M, reports of dead or injured individuals; S, sightings
Discussion
Hunting and incidental drowning of the Antillean manatee in fishing nets in the Orinoco appears to be seasonal. Both occupation of habitat by manatees and the intensity and type of artisanal fishing used in the Orinoco depend on the availability of aquatic macrophytes and fish, respectively. The availability of these resources is strongly influenced by the seasonal variations in water level. Fig. 3 describes the peaks of manatee mortality related to fishing activity, based on the explanations of local fishermen.

Fig. 3 The dynamics of Antillean manatee mortality in the Orinoco River in relation to the seasonality of fishing activities. The cases of manatees hunted or entangled in fishing nets occurred during 1980–2004 (see text for details). The grey area represents the seasonal water level fluctuation. During the low water period (January–April) manatees are isolated in buceaderos (the deepest areas of the river). When the water rises (May–June) manatees migrate in search of food. The peak of the net-fishing season is in the rising water period, which is thus the time of highest entanglement risk for manatees. During the high water period (July–October) manatees occupy the floodplains (esteros). In this season fishing activity is minimal because fish density falls and captures in fishing nets decrease. However, hunting for manatees with harpoons increases. The falling water period starts in November, and the manatees return to the areas of deep water.
In the dry period (January–April) there is little vegetation available for the manatee (Correa-Viana et al., Reference Correa-Viana, O'Shea, Ludlow and Robinson1990) and the water level of the tributaries recedes so much that the manatees are isolated in buceaderos, where they probably feed on dead material from the bottom or algae on rocks. During December–March the river is at its most transparent (Castelblanco-Martínez, 2004); most of the fishing for palometa (Mylossoma spp.) is by hook and the use of nets is infrequent. This would explain the relatively low occurrence of incidental captures in fishing nets during the dry period. Although Mondolfi (Reference Mondolfi1974) reports the hunting of manatees in the dry period, most of the hunters interviewed said that it is not a good period to harpoon the species.
As the river starts to rise (May–June) water covers the beaches and triggers the growth of aquatic macrophytes, which spread rapidly. This is when the manatees start moving to look for food and refuge in the margins of the river. Water transparency decreases drastically, making it impossible to fish with hooks, and nets then become the main means of fishing. During this period nets are put in areas that could be important corridors for manatees, increasing the risk of them being caught.
In the high water period (July–October) fishing activity is minimal because yield in terms of density and size of fish drops markedly (Barbarino Duque & Winemiller, Reference Barbarino Duque and Winemiller2003). In addition, the manatees spread out in the esteros (a vernacular name for the flooded plain of the Orinoco River and tributaries), occupying fields of floating grass or flooded forests, where the use of nets is infrequent and thus incidental captures diminish. However, it is the best period for harpoon hunting, with the largest number of manatees harpooned during this period. O'Shea et al. (Reference O'Shea, Correa-Viana, Ludlow and Robinson1988) also documented a high frequency of manatee hunting in the high water period in the Orinoco. The explanation given by the interviewees is that in the high water period hunting is easier because of the presence of feeding areas; the signs of feeding on the vegetation allow the manatees to be tracked. In addition it is difficult for a manatee to escape if harpooned in the estero because the harpoon usually entangles in the vegetation.
In November the water level begins to recede, the forest dries and the floating communities of macrophytes diminish. As a result, the manatees move from the esteros to the centre of the river, looking for sources of food such as plants on rocks.
The reproductive cycles of the fauna of river systems, including aquatic mammals, are synchronized to the environmental changes caused by the flood pulse (Robinson et al., Reference Robinson, Tockner and Ward2002). During the dry period of 2001 we observed a group of adult manatees in the area of San José. At least five adults were seen to push and bump each other for a 2-hour period, probably carrying out a mating ritual (Hartman, Reference Hartman1979). Local fishermen claimed that mating behaviour of the manatee generally takes place during the dry season. The lower density of manatees during the high water period could reduce the possibility of encounters but in the dry period individuals are confined in relatively limited areas. The differential use of habitats seasonally could thereby determine the seasonality of reproduction (Marmontel et al., Reference Marmontel, Odell, Reynolds and Hemlett1992). Further studies are needed to determine any seasonality in the reproduction of manatees in the Orinoco and the factors regulating the reproductive cycle of this population.
The Florida manatee carries out seasonal migrations, covering hundreds of kilometres (Deutsch et al., Reference Deutsch, Reid, Bonde, Easton, Kochman and O'Shea2003) in response to seasonal variations in water temperature and the necessity to drink fresh water (Hartman, Reference Hartman1979). However, it is probable that the manatees of the Orinoco do not move far (Correa-Viana et al., Reference Correa-Viana, O'Shea, Ludlow and Robinson1990), living permanently in fresh water > 1,100 km from the sea. The habitat of the T. m. manatus population that lives in the Orinoco River can be compared to that of the Amazonian manatee T. inunguis (Junk et al., Reference Junk, Bayley, Sparks and Dodge1989; Colonello, Reference Colonello1990). Both live in dynamic river systems that have high seasonal variations in water levels, and consequently in their depth, current, transparency, margin characteristics, intensity and type of fishing and movement of boats. Such changes determine the availability of habitat for the manatee, and oblige individuals to move locally in search of suitable conditions. Although further information is needed to confirm the hypothesis, evidence suggests a converging adaptation in the behaviour and physiology of the Orinoco and Amazon manatees in response to similar environmental characteristics. It is possible that adaptation to seasonally minimal resources (Colares & Colares, Reference Colares and Colares2002) could be more efficient energetically than long distance migrations (Reid et al., Reference Reid, Rathbun and Wilcox1991; Deutsch et al., Reference Deutsch, Bonde and Reid1998).
Our results indicate that habitat use by the Antillean manatee in the Orinoco depends on the availability of vegetation in the high water period and on the presence of deep-water areas in the dry period. These results also show that the main causes of mortality of the manatee are anthropogenic and influenced by hydrological fluctuations. Fishing activities and movement of boats need to be studied to determine the influence of these factors on reproduction and migration. During the transition periods areas such as Ventanas or Tronconal are important corridors and therefore require appropriate management.
Our results have been taken into account in several manatee conservation initiatives promoted by NGOs and local governments. We further recommend continued educational campaigns with the local communities along the Orinoco River, to discuss the main conservation problems of the manatee and clarify and divulge the existing legislation. The results and conclusions of this study formed part of the basis for the establishment of a national plan for the management and conservation of manatees in Colombia (Caicedo-Herrera et al., 2005), and the management plan for the threatened species of the Reserva de Biósfera El Tuparro (Trujillo et al., Reference Trujillo, Portocarrero and Gómez2008).
Acknowledgements
This study was possible thanks to the logistical support offered by Fundación Omacha and the help of its directors, researchers and drivers. The project received financial help from Sirenian International, Columbus Zoo and Aquarium, Iniciativa para Especies Amenazadas (Convenio Fondo para la Acción Ambiental/Fundación Omacha/Conservación Internacional–Colombia). A study grant was provided by the convention Programa de Estudante-Convênio de Pós-Graduação of Ministério de Ciência e Tecnologia of Brazil. We also recognize the help of Instituto de Ciencias Naturales (Universidad Nacional de Colombia), Universidad Jorge Tadeo Lozano, and Instituto Nacional de Pesquisas da Amazônia (Brazil). Sarita Kendall, Dalila Caicedo, Benjamin Morales-Vela, Vera da Silva and Daryl P. Domning provided insightful comments. Dr Robert Bonde, Mario Rivera and two anonymous referees helped to improve the article. We especially thank the spontaneous collaboration of the fishing communities in the study area. We thank Kesä K. Lehti for the translation.
Biographical sketches
Nataly Castelblanco-Martínez has developed research and educational projects on aquatic fauna in South American rivers, studying the distribution, ecology and conservation of manatees, dolphins, otters and turtles. She is now studying the ecology of Caribbean manatees in Mexico. Isabel Gómez and Lucia Bermudez participated in a manatee conservation project in the Orinoco River for their graduate studies. Fernando C.W. Rosas has been studying the ecology and conservation of aquatic mammals in the Brazilian Amazon for over 25 years. Fernando Trujillo is founder and scientific director of the Fundación Omacha, an NGO created to protect fauna and aquatic ecosystems in Colombia. Enrique Zerda-Ordoñez teaches animal behaviour at the Universidad Nacional de Colombia.








