


Skeletal muscle has a remarkable ability to adapt its metabolism in response to the nutrient intake and energy demands of the organism. In fact, both food deprivation and high-fat diet feeding favour the use of lipids by skeletal muscle as an energy source at the expense of glucose, a phenomenon known as metabolic flexibility. Impaired metabolic flexibility has been shown to be associated with obesity and insulin resistance( Reference Kelley, Goodpaster and Wing 1 , Reference Storlien, Oakes and Kelley 2 ), but the exact mechanisms linking metabolic inflexibility to these pathological conditions are not known. It is also not known whether the lack of adaptation to metabolic demands is a cause or a consequence of obesity or diabetes.
It is now widely accepted that malnutrition during early life is an important predisposing factor for the development of obesity and metabolic diseases. Indeed, based on epidemiological studies demonstrating that people born with a low birth weight are at a high risk of developing the metabolic syndrome in adulthood, Hales & Barker( Reference Hales and Barker 3 ) proposed that in a developing organism exposed to nutrient deficiency, there is a selective distribution of nutrients to preserve brain growth at the expense of other organs. This results in the ‘programming’ of energy metabolism to promote survival under conditions of food restriction. However, when the adverse nutritional environment disappears, the physiological incompatibility between the ‘thrifty’ metabolic programming and the new nutritional conditions leads to obesity, impaired glucose tolerance, hypertension and atherosclerosis( Reference Hales and Barker 3 ).
Experimental studies in different animal species have provided major support for the hypothesis proposed by Hales & Barker( Reference Hales and Barker 3 ) and have led to the identification of several cellular and physiological adaptive responses as potential underlying mechanisms of metabolic programming including alterations in skeletal muscle. Notably, it has been shown that reduced nutrient supply during the last 3 d of gestation in rats leads to decreased ATP production and decreased glucose transport and glycogen biosynthesis in skeletal muscle( Reference Selak, Storey and Peterside 4 ). It has also been shown that offspring born to dams fed a low-protein diet during gestation and lactation initially exhibit better glucose tolerance and enhanced insulin sensitivity in the soleus muscle( Reference Ozanne, Wang and Coleman 5 ), a skeletal muscle containing a high percentage of type I fibres, but become insulin resistant later in life( Reference Ozanne, Olsen and Hansen 6 ). In addition to these functional alterations, a decrease( Reference Mallinson, Sculley and Craigon 7 ), an increase( Reference Toscano, Manhaes-de-Castro and Canon 8 ) or no change( Reference Bayol, Jones and Goldspink 9 ) in the number of type II fibres has been documented in rats following perinatal undernutrition. In contrast to type I fibres, which have high oxidative capacity, type II fibres posses high glycolytic activity and their density is increased in obese and insulin-resistant patients.
The overall objective of the present study was to define the consequences of protein restriction during gestation and lactation on the metabolic flexibility of skeletal muscle in adulthood. Our working hypothesis was that nutritional restriction during the critical period of perinatal development impairs the ability of skeletal muscle to adapt to the use of energy substrates (lipids and glucose) in response to the nutritional and energy demands of the body. Specifically, we examined the influence of protein restriction during gestation and lactation on the expression of a set of genes involved in the starvation-induced transcriptional programme leading to a switch in fuel usage in skeletal muscle from glucose to fat. The analysed genes included the nuclear receptors PPARα and PPARδ and the co-regulator of transcription PPAR-gamma, co-activator 1α (PGC-1α), as well as several genes that play a key role in fatty acid oxidation (carnitine palmitoyltransferase 1a (CPT1a), uncoupling protein 3 (UCP3), pyruvate dehydrogenase kinase, isozyme 4 (PDK4) and medium-chain acyl coenzyme A dehydrogenase (MCAD)). The gene myosin heavy chain class 7 (Myh7), a typical marker of slow-twitch (oxidative) fibres, was also analysed.
Materials and methods
Animals and experimental manipulations
We used a typical model of metabolic programming, which has been described in detail elsewhere( Reference Orozco-Solis, Lopes de Souza and Barbosa Matos 10 ). In brief, virgin female Wistar rats weighing 200–220 g were obtained from Charles River and mated with 3-month-old males of the same strain. After confirmation of mating, pregnant rats were housed individually and fed ad libitum during gestation and lactation either a control diet (170 g protein/kg) or an isoenergetic low-protein diet (80 g protein/kg). The diets were kindly provided by the Department of Nutrition of the Federal University of Pernambuco (Brazil), and their composition has been described previously( Reference Toscano, Manhaes-de-Castro and Canon 8 ). All the experiments were carried out in accordance with the European Communities Council Directive of 24 November 1986 (86/609/EEC) and the Principles of Laboratory Animal Care (NIH publication no. 85–23, revised 1985) and were approved by the Ethical Committee for Animal Experimentation of the Pays de la Loire (protocol CEEA.2012.15). At 1 d after delivery, litter size was adjusted at random to eight pups per dam in both experimental groups maintaining an equal sex ratio. At weaning (21 d), female pups were excluded from the study to yield a final sample size of four male pups per dam. All pups from the control (C) and protein-restricted (PR) groups were fed standard chow until the end of the experiment. The body weight of the pups was recorded every 3 d during lactation and every week after weaning.
Killing and tissue sampling
At 4 months of age, rats were killed under ad libitum feeding conditions or after a fasting period of 48 h. The soleus and extensor digitorum longus (EDL) muscles from both legs were carefully dissected under sodium pentobarbital anaesthesia (50 mg/kg), immediately frozen in liquid N2 and kept at − 80°C until analysis. The rats were then killed by cervical dislocation. All rats were killed within the first hour of the light cycle to avoid the bias induced by the circadian variations in gene expression. Bilateral fat depots from three regions (inguinal, retroperitoneal and epididymal) as well as the mesenteric fat pad were dissected, weighed and pooled to obtain a measure of body fat, referred to as abdominal fat herein. At the end of the experimental period, a total of twelve dams (six controls and six PR) were selected for analysis. From each control and PR litter, two pups were selected at random to obtain a total of twelve control pups and twelve PR pups. From each group, six pups were killed under ad libitum feeding conditions and the other six after fasting. The muscle samples collected from the right and left legs were alternatively used for the determination of gene expression by real-time quantitative PCR and for Western blot analysis. Care was taken to allocate the two pups of the same dam to different experimental groups (fed ad libitum or fasted before killing), such that the presented results corresponded to the combined data from six animals born to six different dams.
Hormone and metabolite measurements
Insulin resistance was evaluated 15 d before killing by the glycaemic response to an oral glucose tolerance test after an overnight fast of 16 h. In brief, a drop of blood was collected immediately before the administration of glucose and 20, 40, 80, and 120 min after administration by severing the tip of the tail. Blood glucose concentrations were determined using a blood glucose monitor (Accu-Check® Active, Roche Diagnostics). To determine the glucose disappearance rate (K G), the AUC was calculated using GraphPad Prism 5.0 (GraphPad Software, Inc.), and the y value of the starting data point was considered to be the baseline value. Plasma from trunk blood collected at the time of killing was assayed for insulin and leptin using assay kits obtained from Millipore. Enzymatic diagnostic kits obtained from BioMérieux were used to quantify the plasma concentrations of TAG, glucose and cholesterol. The concentrations of NEFA were determined using an assay kit obtained from Wako.
Real-time quantitative PCR
Total RNA from the soleus and EDL muscles was extracted using the TRIzol reagent (Invitrogen) and treated with DNAse (RQ1 DNase, Promega) for 45 min at 37°C. Later, 1 μg of purified RNA was reverse-transcribed using M-MLV Reverse Transcriptase (Promega) in a total volume of 25 μl, and the resulting complementary DNA was diluted 40-fold in DNAse and RNAse-free water. Thereafter, 5 μl of each complementary DNA diluted sample were used as a template for PCR amplification using SYBR Green (Qiagen) as a fluorogenic intercalating dye and the iCycler iQ Real-Time PCR Detection System from Bio-Rad Laboratories. The PCR parameters were as follows: an initial denaturation step of 5 min at 95°C, followed by forty cycles of 30 s at 95°C and 30 s at 60°C.
Differences in gene expression among the groups were determined using the comparative threshold cycle (C
t) method as described by Schmittgen & Livak(11) and 18S RNA as the housekeeping gene. In brief, the C
t value was automatically determined for all the targeted genes and 18S in all the samples and within the same experiment. The relative amount of the different genes normalised to the endogenous expression of 18S was calculated using the following formula:
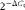 $$2^{ - \Delta C _{t}} $$
, where ΔC
t= C
t(gene)− C
t(18S). Fold changes in gene expression due to the different nutritional manipulations were then calculated by dividing the
$$2^{ - \Delta C _{t}} $$
value of each sample by the averaged
$$2^{ - \Delta C _{t}} $$
value of the control group. Primers were designed using the information stored in the public GenBank database of the National Center for Biotechnology Information, and their sequences are given in Table 1. The applicability of the C
t method and the use of 18S as the housekeeping gene were first validated by determining how the amplification efficiencies of the different transcripts in relation to those of 18S, β-actin and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) varied with template dilution. These preliminary experiments indicated that the PCR gave a single product of the expected size for all the genes, as determined by the melting curve analysis and agarose gel electrophoresis (data not shown), and that the amplification efficiency of the target gene was the same independently of the use of 18S, β-actin or GAPDH as a calibrator (Fig. S1(A), available online). It was then verified whether the expression of 18S as well as that of β-actin or GAPDH was altered by fasting or by the early exposure to protein restriction (Fig. S1(B), available online). Finally, as shown in Fig. S1(C) (available online), it was confirmed that the pattern of gene expression in the different groups of rats was the same independently of the housekeeping gene used.
$$2^{ - \Delta C _{t}} $$
, where ΔC
t= C
t(gene)− C
t(18S). Fold changes in gene expression due to the different nutritional manipulations were then calculated by dividing the
$$2^{ - \Delta C _{t}} $$
value of each sample by the averaged
$$2^{ - \Delta C _{t}} $$
value of the control group. Primers were designed using the information stored in the public GenBank database of the National Center for Biotechnology Information, and their sequences are given in Table 1. The applicability of the C
t method and the use of 18S as the housekeeping gene were first validated by determining how the amplification efficiencies of the different transcripts in relation to those of 18S, β-actin and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) varied with template dilution. These preliminary experiments indicated that the PCR gave a single product of the expected size for all the genes, as determined by the melting curve analysis and agarose gel electrophoresis (data not shown), and that the amplification efficiency of the target gene was the same independently of the use of 18S, β-actin or GAPDH as a calibrator (Fig. S1(A), available online). It was then verified whether the expression of 18S as well as that of β-actin or GAPDH was altered by fasting or by the early exposure to protein restriction (Fig. S1(B), available online). Finally, as shown in Fig. S1(C) (available online), it was confirmed that the pattern of gene expression in the different groups of rats was the same independently of the housekeeping gene used.
Table 1 Sequences of primers used in the real-time RT-PCR analysis

Western blot analysis
Muscle samples were homogenised in 1:8 volume (w/v) of extraction buffer containing 50 mm-HEPES (pH 7·6), 50 mm-KCl, 50 mm-NaF, 5 mm-Na4P2O7, 1 mm-EDTA, 1 mm-ethylene glycol tetraacetic acid, 1 mm-dithiothreitol, 5 mm-β-glycerophosphate, 1 % NP-40, 1 mm-Na3VO4 and a cocktail of protease inhibitors (Sigma, P2714). Homogenates were then centrifuged at 10 000 g at 4°C for 10 min. The resulting supernatants were used for Western blot analysis. In brief, 100 μg of protein extracts were separated on a 10 % SDS–PAGE gel and transferred onto polyvinylidene difluoride membranes. Non-specific binding was blocked by incubating the membranes for 1 h at room temperature with 5 % bovine serum albumin dissolved in Tris-buffered saline with Tween-20 (TBST, 10 mm-Tris–HCl, 150 mm-NaCl and 0·1 % Tween 20; pH 7·5). The membranes were then incubated overnight at 4°C with a primary antibody against total AMP-activated protein kinase (AMPK) or with a rabbit polyclonal antibody recognising the phosphorylated form of AMPK at Thr172. Both antibodies were used at a dilution of 1:1000 and were purchased from Cell Signaling. To control for protein loading, the same blots were incubated with an anti-α-tubulin antibody (Santa Cruz Biotechnology, Inc.) diluted at 1:100. Later, the membranes were incubated for 1 h at room temperature with goat anti-rabbit peroxidase-conjugated secondary antibodies (Jackson ImmunoResearch). Immunoblotted proteins were visualised using the UptiLight™ HRP blot substrate (Interchim) and analysed using the G:BOX Chemi XL system (Syngene).
Data analysis
Experimental results are expressed as means with their standard errors. Statistical differences in gene expression under ad libitum feeding conditions between the control and PR rats were assessed using Student's t test. To determine the effects of fasting on plasma metabolite concentrations and gene expression, two-way ANOVA was used followed by Bonferroni's test with maternal diet (control or PR) and nutritional status at killing (ad libitum or fasted) as independent factors. Body weight gain was calculated using the following formula: ((W n+1− W n /W n )) × 100, where W n is weight in g on day ‘n’ and W n+1 is weight in g on the next recording day. Differences in body weight were assessed with two-way repeated-measures ANOVA using maternal diet and age as factors. Statistical significance was set at P< 0·05.
Results
Phenotypic characteristics of the pups and metabolite variations in plasma in response to fasting
There were no differences in body weight at birth between control and PR pups (C = 5·21 (sem 0·18) g, n 12; PR = 4·86 (sem 0·21) g, n 12). However, the offspring born to, and nursed by, PR dams gained significantly less weight during lactation such that at the end of this period they weighed, on average, 58 % less than their control counterparts (C = 43·38 (sem 0·46) g, n 12; PR = 19·50 (sem 0·66) g, n 12, P< 0·0001; Fig. 1(a)). During the first 2 months after weaning, the PR pups exhibited enhanced body weight gain when compared with control pups (Fig. 1(b)). Nevertheless, in spite of this catch-up growth, they continued to weigh less than control pups until the end of the experimental period (Fig. 1(a)). In addition, the PR pups exhibited less abdominal fat both in absolute terms (C = 18·88 (sem 2·01) g, n 12; PR = 11·48. (sem 1·32) g, n 12; P< 0·01) and in proportion to their body weight (C = 8·26 (sem 0·70) %, n 12; PR = 4·82 (sem 0·32) %, n 12; P< 0·001).

Fig. 1 Evolution of (a) body weight and (b) body weight gain in male rats born to dams fed a control diet (17 % protein) or a low-protein diet (8 % protein) during gestation and lactation. At weaning, all rats were fed standard chow. Data in (b) correspond to the percentage in body weight gain between the indicated recording periods. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the control group: **P< 0·01; ***P< 0·0001 (two-way repeated-measures ANOVA). ○, Control; ●, protein restricted.
Control and PR rats exhibited the same glucose tolerance as indicated by the identical glucose disappearance rates between 20 and 120 min after the oral administration of glucose (C = 308·20 (sem 32·53) mmol/l × 120 min, n 12; PR = 309·80 (sem 41·54) mmol/l × 120 min, n 12) and the same temporal decrease in plasma glucose concentrations after the glucose load (data not shown). With the exception of a reduced concentration of TAG and insulin, the plasma metabolite profile of the control and PR adult rats was the same under ad libitum feeding conditions (Fig. 2). Both groups also exhibited similar variations in plasma metabolite concentrations in response to a 48 h fast. Notably, however, fasting reduced the plasma content of insulin in control rats but not in PR rats (Fig. 2).

Fig. 2 Metabolite concentrations in the plasma of adult rats born to dams fed a control diet (17 % protein) or a low-protein diet (8 % protein) during gestation and lactation. Rats were killed under ad libitum feeding conditions within the first hour of the light cycle or after a 48 h fast. Data were analysed using two-way ANOVA using maternal diet (control or protein restricted) and nutritional status at killing (ad libitum (□) or fasted (![]() )) as independent factors followed by Bonferroni's test. (a) Glucose, (b) cholesterol, (c) TAG, (d) NEFA, (e) leptin and (f) insulin. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the ad libitum-fed animals of the same group: * P< 0·05, ** P< 0·01, *** P< 0·001. ††† Mean value was significantly different from that of the ad libitum-fed control group animals (P< 0·001).
)) as independent factors followed by Bonferroni's test. (a) Glucose, (b) cholesterol, (c) TAG, (d) NEFA, (e) leptin and (f) insulin. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the ad libitum-fed animals of the same group: * P< 0·05, ** P< 0·01, *** P< 0·001. ††† Mean value was significantly different from that of the ad libitum-fed control group animals (P< 0·001).
Gene expression profile under ad libitum feeding conditions
The mRNA expression of several genes involved in the regulation of fatty acid oxidation was higher in soleus muscle, but not in EDL muscle, of PR rats killed in the fed state than in the muscles of their control counterparts. Thus, the mRNA expression of CPT1a and MCAD was up-regulated in PR rats (Fig. 3). CPT1a is the key enzyme in the carnitine-dependent transport of fatty acids across the mitochondrial inner membrane and MCAD catalyses the first dehydrogenation step in the β-oxidation of fatty acids. Moreover, the expression of the nuclear receptor PPARα as well as that of the co-regulator of transcription PGC-1α, which in a coordinated fashion positively regulate the expression of several genes playing a key role in fatty acid oxidation, was increased by more than 2-fold by perinatal protein restriction (Fig. 3). Similar differences in the expression of mRNA encoding the α2 isoform of AMPK were observed between control and PR rats. This kinase is activated by energy depletion and regulates a large number of cellular process that synergistically inhibit protein and lipid biosynthesis and stimulate β-oxidation, glucose uptake and glycolysis( Reference Lage, Dieguez and Vidal-Puig 12 ). Finally, using Student's t test, an increased expression of UCP3, which exports NEFA from the mitochondrial matrix when fatty acid supply exceeds oxidation( Reference Abe, Mujahid and Sato 13 , Reference Himms-Hagen and Harper 14 ), and a decreased expression of PDK4, which negatively regulates glucose metabolism through the inhibition of the pyruvate dehydrogenase complex, were also detected (Fig. 3). In contrast to these multiple gene expression changes in soleus muscle, the only transcriptional change detected in EDL muscle of ad libitum-fed PR rats was an increased expression of mRNA encoding PPARα and PPARδ nuclear receptors along with a decreased expression of MCAD (Fig. 4).

Fig. 3 Gene expression changes in the soleus from control and protein-restricted rats in response to fasting: (a) carnitine palmitoyltransferase 1a; (b) medium-chain acyl coenzyme A dehydrogenase; (c) β-hydroxyacyl-coenzyme A dehydrogenase; (d) uncoupling protein 3; (e) pyruvate dehydrogenase kinase, isozyme 4; (f) PPAR-gamma, co-activator 1α; (g) PPARα; (h) PPARδ; (i) AMP-activated protein kinase α2; (j) myosin heavy chain class 7. Variations in gene expression are illustrated in relation to those observed under ad libitum feeding conditions using the expression in fed animals of the control group as a calibrator. (□), Ad libitum; (![]() ), fasted. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the ad libitum-fed animals of the same group: * P< 0·05, ** P< 0·01, *** P< 0·001 (two-way ANOVA followed by Bonferroni's test). † Mean value was significantly different from that of the ad libitum-fed control group animals (P< 0·05; Student's t test).
), fasted. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the ad libitum-fed animals of the same group: * P< 0·05, ** P< 0·01, *** P< 0·001 (two-way ANOVA followed by Bonferroni's test). † Mean value was significantly different from that of the ad libitum-fed control group animals (P< 0·05; Student's t test).

Fig. 4 Gene expression changes in the extensor digitorum longus from control and protein-restricted rats in response to fasting: (a) carnitine palmitoyltransferase 1a; (b) medium-chain acyl coenzyme A dehydrogenase; (c) β-hydroxyacyl-coenzyme A dehydrogenase; (d) uncoupling protein 3; (e) pyruvate dehydrogenase kinase, isozyme 4; (f) PPAR-gamma, co-activator 1α; (g) PPARα; (h) PPARδ; (i) AMP-activated protein kinase α2; (j) myosin heavy chain class 7. Variations in gene expression are illustrated in relation to those observed under ad libitum feeding conditions using the expression in fed animals of the control group as a calibrator. (□), Ad libitum; (![]() ), fasted. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the ad libitum-fed animals of the same group: * P< 0·05, ** P< 0·01, *** P< 0·001 (two-way ANOVA followed by Bonferroni's test). † Mean value was significantly different from that of the ad libitum-fed control group animals (P< 0·05; Student's t test). ‡‡ Mean value was significantly different from that of the fasted control group animals (P< 0·01).
), fasted. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the ad libitum-fed animals of the same group: * P< 0·05, ** P< 0·01, *** P< 0·001 (two-way ANOVA followed by Bonferroni's test). † Mean value was significantly different from that of the ad libitum-fed control group animals (P< 0·05; Student's t test). ‡‡ Mean value was significantly different from that of the fasted control group animals (P< 0·01).
Gene expression changes in response to fasting
Both divergent and similar mRNA transcription changes were observed in response to fasting in control and PR rats and these changes differed from one muscle type to the other. Thus, while fasting increased the expression of CPT1a in soleus muscle of control rats, no change in mRNA expression was detected in fasted PR rats when compared with ad libitum-fed PR rats (Fig. 3). Moreover, fasting decreased the expression of MCAD and β-hydroxyacyl coenzyme A dehydrogenase (β-HAD) and AMPKα2 in PR rats but not in control rats (Fig. 3). Similar to MCAD, β-HAD catalyses several steps of the β-oxidation of long-chain fatty acids in the mitochondria. In addition, the expression of PDK4 was not affected by fasting neither in control nor PR rats (Fig. 3). Among the genes involved in the transcriptional regulation of metabolism (PGC-1α, PPARα and PPARδ), the expression of myosin heavy chain 7 (Myh7), which defines the formation of type I (oxidative) fibres, was decreased by fasting in both control and PR rats (Fig. 3). However, it should be noted that in the case of PGC-1α and PPARα, the differences in gene expression between the ad libitum-fed and fasted control rats reached statistical significance only when the data were analysed using Student's t test.
With regard to the transcriptional changes in EDL muscle, it was found that fasting increased the expression of CPT1a and UCP3 in both groups of rats (Fig. 4). However, fasted PR, but not fasted control rats, exhibited an increased expression of PDK4 and a decreased expression of mRNA encoding PPARδ in relation to their ad libitum-fed counterparts (Fig. 4). Moreover, the expression of MCAD was down-regulated by fasting in the control but not in PR rats. Lastly, food deprivation for 48 h did not alter the mRNA expression of β-HAD, PGC-1α, AMPKα2 and MHC1 neither in control nor in PR rats (Fig. 4).
Effect of maternal protein restriction on the activation of AMP-activated protein kinase
The nutrient sensor AMPK is activated by energy depletion and once activated regulates a wide range of cellular processes that synergistically inhibit protein and lipid biosynthesis and stimulate β-oxidation, glucose uptake and glycolysis. Therefore, it was surprising to find no changes in the expression of AMPKα2 in the soleus muscle of control fasted rats. To clearly determine whether AMPK was activated under our experimental conditions, the phosphorylated (activated) form of AMPK at Thr172 was quantified by Western blot analysis. As shown in Fig. 5, PR rats exhibited enhanced phosphorylation levels of AMPK in the fed state in soleus but not in EDL. Moreover, whereas fasting induced a significant activation of AMPK in the soleus muscle of control rats, no change in AMPK phosphorylation levels in response to fasting was detected in PR rats (Fig. 5(a)). Fasting also increased the phosphorylation levels of AMPK in the EDL muscle of control and PR rats, though in control rats this change did not reach statistical significance (Fig. 5(b)).

Fig. 5 Changes in AMP-activated protein kinase (AMPK) activity in response to fasting in the (a) soleus muscle and (b) extensor digitorum longus muscle of adult rats born to control or protein-restricted dams. Each bar corresponds to the ratio of phosphorylated levels:non-phosphorylated levels of AMPK determined by Western blot analysis. (□), Data obtained from rats killed under ad libitum feeding conditions. (![]() ), Data obtained from rats killed after a fasting period of 48 h. Mean value was significantly different from that of the ad libitum-fed animals of the same group: * P< 0·05, ** P< 0·01 (two-way ANOVA followed by Bonferroni's test). † Mean value was significantly different from that of the ad libitum-fed control group animals (P< 0·05; two-way ANOVA followed by Bonferroni's test). pAMPK, phospho AMPK; AU, arbitrary units.
), Data obtained from rats killed after a fasting period of 48 h. Mean value was significantly different from that of the ad libitum-fed animals of the same group: * P< 0·05, ** P< 0·01 (two-way ANOVA followed by Bonferroni's test). † Mean value was significantly different from that of the ad libitum-fed control group animals (P< 0·05; two-way ANOVA followed by Bonferroni's test). pAMPK, phospho AMPK; AU, arbitrary units.
Discussion
While reduced energy expenditure and increased consumption of highly energetic foods are important factors that contributing to the current epidemic of obesity, many epidemiological and experimental studies have demonstrated that an unbalanced provision of nutrients during perinatal development increases the risk of developing obesity and the metabolic syndrome in adulthood( Reference Ozanne and Constancia 15 ). Several mechanisms have been proposed to account for this enhanced disease risk including altered organ structure( Reference Merezak, Reusens and Renard 16 , Reference Vehaskari, Aviles and Manning 17 ), elevated glucocorticoid levels and endocrine sensitivity( Reference Cottrell, Holmes and Livingstone 18 , Reference Fowden and Forhead 19 ), impaired mitochondrial function( Reference Reusens, Theys and Remacle 20 , Reference Leduc, Levy and Bouity-Voubou 21 ) and altered feeding behaviour( Reference Vickers, Breier and Cutfield 22 , Reference Bellinger and Langley-Evans 23 ). In the present study, we showed that the metabolic flexibility of skeletal muscle is impaired in adult offspring born to dams fed a low-protein diet during gestation and lactation adding to the detrimental effects of malnutrition during early life.
Skeletal muscle is a heterogeneous tissue that has different types of fibres, namely type I and type II, which differ in acid stability of their myofibrillar ATPase activity and their contractile and metabolic properties( Reference Punkt, Naupert and Asmussen 24 , Reference Schiaffino and Reggiani 25 ). Type I fibres mainly use fat as an energy substrate, their speed of contraction is relatively slow and they are resistant to fatigue. In contrast, type II fibres use glucose as an energy source, contract quickly and do not have a high resistance to fatigue. On the basis of their pH liability, type II fibres are further categorised into type IIa, which possess a mixture of glycolytic and oxidative properties, and type IIb, which are exclusively glycolytic( Reference Brooke and Kaiser 26 ). The soleus muscle consists of only type I and type IIa fibres, whereas the EDL muscle mainly consists of type IIa and type IIb fibres. Consequently, because of its high oxidative capacity, the soleus muscle tends to use NEFA as energy substrates, whereas the EDL muscle has far higher basal rates of glycolysis and gluconeogenesis. Moreover, these two muscles use different mechanisms to cope with starvation-induced elevated NEFA concentrations. In fact, the soleus muscle disposes of excess NEFA by oxidation, whereas the EDL muscle neutralises NEFA by re-esterification( Reference Neumann-Haefelin, Beha and Kuhlmann 27 ). Therefore, to determine the effects of early malnutrition on the metabolic flexibility of skeletal muscle, we investigated the expression of several genes involved in lipid metabolism and mitochondrial function in the soleus and EDL muscles as they possess opposite metabolic properties.
Under ad libitum feeding conditions, PR rats exhibited increased AMPK gene expression and phosphorylation levels in soleus muscle along with an enhanced mRNA expression of several genes encoding key proteins favouring fatty acid oxidation including CPT1a, PGC-1α, UCP3 and PPARα. In contrast, there was an increased mRNA expression of PPARα and PPARδ along with a decreased gene expression of MCAD in EDL muscle, indicating that, in addition to having divergent metabolic properties, these two muscles are differentially affected by metabolic programming.
Increased β-oxidation( Reference Perdomo, Commerford and Richard 28 ) and overexpression of CPT1 ( Reference Bruce, Hoy and Turner 29 ) in skeletal muscle cells have been shown to improve insulin resistance. The gene expression data obtained in the present study, along with the observation that PR rats also had reduced body weight and decreased plasma concentrations of TAG, indicate that early protein restriction induces a long-lasting metabolic state favouring lipid catabolism and insulin function. This observation seems to be at odds with the thrifty phenotype hypothesis stating that a poor nutritional environment during early life increases the risk of developing the metabolic syndrome in adulthood( Reference Hales and Barker 3 ). However, enhanced capacity for fat oxidation in muscle is not in itself predictive of a healthy phenotype. In fact, transgenic mice in which increased fatty acid oxidation is induced by muscle-specific overexpression of PPARα or PGC-1α are more susceptible to diet-induced obesity and diabetes( Reference Choi, Befroy and Codella 30 ). Moreover, high-fat diet feeding increases the lipid oxidation capacity of skeletal muscle through the up-regulation of several key genes regulating mitochondrial biogenesis( Reference Turner, Bruce and Beale 31 ). On the basis of these results, it has been suggested that incomplete fatty acid oxidation and accumulation of β-oxidative intermediates resulting from the mismatch between lipid overload and energy needs are the primary biochemical defects underlying insulin resistance( Reference Kelley, Goodpaster and Wing 1 , Reference Muoio Deborah and Neufer 32 ). In this respect, it is interesting to note that ad libitum-fed PR rats exhibited an enhanced expression of CPT1a and UCP3 in the soleus muscle without any modification in the expression of mRNA encoding β-HAD. This transcriptional profile is indicative of an enhanced flux of lipids into the mitochondria without a concomitant increase in fatty acid oxidation. This would lead to increased export of fatty acid anions outside the mitochondrial matrix, which might result in the accumulation of lipids in skeletal muscle.
Fasting is a nutritional challenge that leads to rapid metabolic adaptations of skeletal muscle that result in the shift from glucose to fatty acid oxidation as the source of fuel. This phenomenon involves a complex mechanism of transcriptional regulation that includes the nuclear receptors PPARα and PPARδ, the co-transcriptional regulator PGC-1α and the nutrient sensors sirtuin 1 and AMPK( Reference Bassel-Duby and Olson 33 – Reference Rodgers, Lerin and Gerhart-Hines 35 ). The two latter factors, which are activated under physiological conditions that require a large amount of energy such as nutritional restriction or exercise, induce the expression of PGC-1α and directly enhance its activity through deacetylation (sirtuin 1) or phosphorylation (AMPK)( Reference Cantó and Auwerx 36 ). The expression of PGC-1α can also be enhanced via the nuclear receptor PPARδ( Reference Schuler, Ali and Chambon 37 ) and a Ca-dependent signalling pathway involving the activation of the transcription factor C-AMP response element-binding protein (CREB)( Reference Chowanadisai, Bauerly and Tchaparian 38 ). Once activated, PGC-1α facilitates the expression of several genes involved in fatty acid oxidation and mitochondrial function by forming a complex with the nuclear receptor PPARα( Reference Vega, Huss and Kelly 39 ) and, simultaneously, regulates the reprogramming of fast-twitch, glycolytic fibres (type II) into slow-twitch, oxidative fibres (type I) through the co-activation of myosin enhancer factor 2( Reference Lin, Wu and Tarr 40 ).
The transcriptional responses of skeletal muscle to fasting are time dependent. Thus, the expression of PPARδ and PGC-1α is initially up-regulated by food deprivation, but after a 48 h fast, it decreases below control levels( Reference de Lange, Farina and Moreno 41 ). In agreement with these results, we observed that a 48 h starvation period reduced the expression of these two transcription factors as well as that of PPARα in the soleus muscle of both control and PR rats. However, we found a clear difference in the gene expression profiles of CPT1a, MCAD and β-HAD between control and PR rats in response to fasting. In fact, the mRNA levels of the former gene were increased by starvation in control rats but not in PR rats. Moreover, whereas fasted PR rats exhibited a reduced expression of MCAD and β-HAD in relation to their ad libitum-fed counterparts, starvation induced no change in the expression of these genes in control rats. Similarly, the activation of AMPK, as measured by the phosphorylation of its α-subunit at Thr172, was increased by fasting in control but not in PR rats. Altogether, these gene expression changes are indicative of impaired fatty acid uptake and oxidation in response to fasting in the soleus muscle of PR rats. Furthermore, as expected from their divergent metabolic properties, the soleus and EDL muscles reacted differently to food deprivation and their metabolic flexibility was differentially affected by early protein restriction. Thus, while starvation down-regulated the expression of the transcriptional regulators of lipid metabolism (PPARα, PPARδ and PGC-1α) in the soleus muscle of both control and PR rats, this effect was observed only in the EDL muscle of PR rats. In addition, in sharp contrast to its effects in soleus muscle, fasting induced a gene expression profile indicative of enhanced fatty acid β-oxidation and reduced glycolysis in EDL muscle of PR rats. Collectively, these results show that fasting alters the metabolic properties of skeletal muscle in a very fine and selective manner.
Kelley et al. ( Reference Kelley, Goodpaster and Wing 1 ) were the first to propose that the metabolic inflexibility of skeletal muscle, defined as decreased fatty acid oxidation under fasting conditions or reduced capacity to switch from lipid oxidation to glucose uptake and catabolism during the transition from fasting to feeding, is associated with obesity and diabetes and predicts the severity of insulin resistance. Further studies have corroborated that skeletal muscle of obese or diabetic patients exhibits decreased fatty acid oxidation during fasting or exercise as well as reduced capacity to shift between lipid and glucose oxidation under insulin-stimulating conditions( Reference Blaak, van Aggel-Leijssen and Wagenmakers 42 , Reference Thyfault, Rector and Noland 43 ). However, it has not been determined whether metabolic inflexibility is an additional manifestation of obesity and diabetes or whether it is an underlying cause of these metabolic disorders. In the present study, we showed that 4-month-old rats born to PR dams exhibit an altered gene expression pattern in skeletal muscle in response to fasting. The fact that, at this age, these animals did not exhibit any sign of obesity or glucose intolerance suggests that metabolic inflexibility might be the cause and not the consequence of obesity and diabetes. Interestingly, early protein restriction differentially affected the responses of the soleus and EDL muscles to fasting, further indicating that the metabolic inflexibility of skeletal muscle might be fibre type specific.
In summary, the results of the present study indicate that under ad libitum feeding conditions adult offspring born to PR dams exhibit a gene expression pattern in skeletal muscle favouring the transport of long-chain fatty acids into the mitochondrial matrix without a concomitant increase in fatty acid oxidation. Moreover, the transcriptional network regulating the metabolic switch from carbohydrate to lipid oxidation during fasting was clearly altered in the PR rats, which otherwise exhibit a normal metabolic state. This observation suggests that impaired metabolic flexibility precedes and may underpin the development of obesity and insulin resistance induced by maternal protein restriction. Given that the deleterious effects of early malnutrition develop with age, further studies are warranted to determine how the herein reported impaired metabolic plasticity of skeletal muscle might contribute to insulin resistance and obesity.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0007114514000865
Acknowledgements
The authors cordially thank Dr Elizabeth do Nascimento from the Nutrition Department of the Federal University of Pernambuco for preparing the experimental diets.
R. d. S. A. was the recipient of a doctoral fellowship from the Brazilian Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). O. G.-Q. and G. P.-G. were the recipients of, respectively, a doctoral and a post-doctoral fellowship from the Mexican Consejo Nacional de Ciencia y Tecnología. The present study was partially supported by CAPES-COFECUB (Action Number Me 657/09). The funders had no role in the design and analysis of the study or in the writing of this article.
The authors' contributions are as follows: F. B.-J. conceived the study, designed the experiments, analysed the data and wrote the manuscript; R. d. S. A. carried out the experiments and contributed to the writing of the manuscript; O. G.-Q. and G. P.-G. conducted the experiments; R. M.-d.-C. contributed to the design of the experiments.
The authors have no competing interests to declare.







