


Obesity is the most common nutritional disorder in the developed world and is a strong risk factor for hypertension, hyperlipidaemia, CVD and type 2 diabetes mellitus, which are closely linked with insulin resistance, and collectively called the metabolic syndromeReference Spiegelman and Flier1. Obesity causes excess fat accumulation not only in adipocytes but also ectopically in tissues such as muscle, liver, β cells and others, predisposing to the development of insulin resistance. Especially, skeletal muscle is a major site for insulin-stimulated glucose disposalReference Boden, Lebed, Schatz, Homko and Lemieux2 and the accumulation of TAG within lipid droplets in skeletal muscle is positively correlated to the severity of insulin resistanceReference Forouhi, Jenkinson, Thomas, Mullick, Mierisova, Bhonsle, McKeigue and Bell3, Reference Krssak, Falk Petersen, Dresner, DiPietro, Vogel, Rothman, Roden and Shulman4.
Recently, there has been growing interest in the use of medical plants and health foods for the treatment and prevention of diseaseReference Crespy and Williamson5, Reference Wang and McIntosh6. Therefore, studies on obesity and diabetes as lifestyle-related diseases have focused on the search of functional food ingredients that suppress the accumulation of body fat and improve lipid metabolismReference Torres, Torre-Villalvazo and Tovar7–Reference Leterme9, effects that, in turn, are beneficial for the amelioration of insulin resistance and prevention of type 2 diabetes.
The chickpea (Cicer arietinum L.) is one of the world's most important legume crops as it contains approximately 50 % available carbohydrate, primarily in the form of starch, and 6·4 % fat, of which most is unsaturated (for example, the fatty acids in the Kabuli-type seeds include 50·3 % oleic acid and 40 % linoleic acid)Reference Thorne, Thompson and Jenkins10. Moreover, the chickpea is considered to be a good source of proteinReference Khan, Akhtar, Ullah and Jaffery11. In addition to culinary usage, the chickpea has been accepted as a natural Uighur traditional medicine in Xinjiang (China) for over 2500 years. As recorded in the pharmacopoeia12, the chickpea is effective in improving plasma lipid levels. Chickpeas may also be beneficial in the treatment of type 2 diabetes because it has a low glycaemic index, indicating that consuming an equi-available carbohydrate portion of chickpeas results in a lower postprandial blood glucose response than standard white wheatflour bread, through having higher levels of slowly digestible starchReference Leterme9, Reference Hawkins and Johnson13, Reference Foster-Powell, Holt and Brand-Miller14.
However, the evidence for the use of chickpeas in insulin resistance and diabetes treatment is few and sometimes discrepantReference Nestel, Cehun and Chronopoulos15–Reference Zulet, Macarulla, Portillo, Noel-Suberville, Higueret and Martinez18. The present study was carried out to further evaluate the effect of chickpeas on modifying lipids, reducing visceral adiposity and aimed at identifying the potential use of chickpeas for the prevention and treatment of obesity, dyslipidaemia and diabetes.
Materials and methods
Materials
Chickpeas were obtained from Xinjiang Amina Agriculture Ltd (Xinjiang, China); rats and specific laboratory diets were purchased from SLAC Laboratory Animal Co. Ltd (Shanghai, China). Other chemicals, solvents and reagents were commercially available.
Animals, diets, and experimental design
Male Sprague–Dawley rats (8 weeks old, about 230 g body weight) were maintained in a temperature-controlled room (22°C) on a 12 h light–dark cycle, and provided with a normal-fat diet (NFD) for 2 weeks after initiation of preliminary rearing; thereafter they were divided into different groups, with the groups matched for body weight. The animals had free access to water and the assigned diets for the different dietary formulations. Three kinds of diets were given in the experiments as follows: (1) a standard laboratory chow (NFD) containing 5 g fat, 22 g protein and 1381 kJ/100 g diet; (2) a high-fat diet (HFD) created by mixing lard (20 %, w/w), sugar (4 %, w/w), whole-milk powder (2 %, w/w) and cholesterol (1 %, w/w) into the standard laboratory chow, which contained 25·71 g fat, 19·54 g protein and 1987 kJ/100 g diet; (3) a high-fat plus chickpea diet (HFD+CP) was the same as the HFD diet, except that 10·0 % of raw crushed chickpea seed replaced the standard laboratory chow, which contained 25·11 g fat, 19·36 g protein and 1965 kJ/100 g diet. The experimental diets were freshly prepared once per month, and stored at − 20°C to avoid rancidity.
Rats (n 30) were randomly assigned to one of the following diets for 8 months: NFD, HFD or HFD+CP. Food intake was monitored over the course of 24 h for 7 consecutive days during the 8-week dietary treatment period, and body weight was measured monthly. At 7 months of diet supplementation, an intraperitoneal insulin tolerance test (ITT; insulin at 2 U/kg) and an oral glucose tolerance test (glucose at 2 g/kg) were performed respectively after an overnight fast, blood glucose levels measured from the tail vein were determined using a glucometer freestyle (in ITT) and the oxidase enzymic method (in the oral glucose tolerance test). At the end of the experiment (8 months after treatment), rats were deprived of food for 12 h and then killed by decapitation between 09.00 and 11.00 hours. The liver, muscle and adipose tissues were removed according to defined anatomical landmarks, the weights of the epididymal adipose tissues were measured, and then were immediately frozen using liquid N2 and kept at − 80°C until use. The serum was separated from blood samples and immediately stored at − 80°C.
Analytic methods
Blood chemistry and hormone analysis
The serum TAG, total cholesterol, LDL-cholesterol (LDL-C), HDL-cholesterol (HDL-C), alanine aminotransferase, blood urea N, creatinine and uric acid levels were measured using Beckman test kits (Beckman Coulter, Inc., Fullerton, CA, USA). Serum insulin and leptin levels were measured using commercial assay kits (rat insulin ELISA kit (Sibayagi, Gunma, Japan) and rat leptin RIA kit (Linco Research Inc., St Charles, MO, USA)).
Measurement of tissue triacylglycerol content
The rats' freeze-dried muscle was powdered, and cleaned of all visible connective tissue and blood under magnification. Lipid was extracted by a Folch extraction as previously describedReference Frayn and Maycock19, the TAG was saponified in an ethanol–KOH solution at 60°C, and glycerol content was determined fluorometrically.
Enzyme assays
The activity of lipoprotein lipase (LPL) in epididymal adipose tissue as well as hepatic TAG lipase (HTGL) in liver were analysed using a non-radioisotopic method according to the kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China), in accordance with the same principle as described previouslyReference Nozaki, Kubo, Matsuzawa and Tarui20. Tissue samples were homogenised in 1 ml of a solution containing 0·25 m-sucrose, 1 mm-EDTA, 10 mm-tri(hydroxymethyl)-aminomethane-HCl and 12 mm-deoxycholate (pH 7·4). The homogenates were centrifuged for 15 min at 3000 rpm (4°C), after which the intermediate phase (between the floating fat droplets and the pellet) was extracted and then incubated under gentle agitation for 1 h at 37°C with a substrate mixture (supplied in the kit). A selective assay of each lipase was performed by addition of SDS and NaCl to inactivate HTGL and LPL, respectively. NEFA were measured by spectrophotometry using the complex cupric acetate-pyridine procedure. LPL and HTGL activity were expressed as microunits (1 μU = 1 μmol NEFA released per h incubation at 37°C).
Extraction and Northern blot analysis of total RNA
7w>Epididymal adipose tissue total RNA was extracted using a TRIZOL reagent (Invitrogen Life Technologies Inc., Gaithersburg, MD, USA). The extracted RNA samples (20 μg) were subjected to 1 % agarose gel electrophoresis in the presence of formalin and then transferred onto a nylon membrane (Hybond-N; Amersham, Little Chalfont, Bucks, UK). Prehybridisation and hybridisation of the blots were performed as described previouslyReference Neschen, Moore, Regittnig, Yu, Wang, Pypaert, Petersen and Shulman21 using specific cDNA probes labelled with Reference McGarry32P dCTP by random priming. The following primers were used: leptin, 5′-TCCCACCACAACGAAGTAC-3′ and 5′-TCAGCCAGGGCATTATTTTC-3′; LPL, 5′-GCTTCACCCCATTCTGAGTT-3′ and 5′-TCCAGGCTCTCTGGCTTCTG-3′; β-actin, 5′-GCCAACCGTGAAAAGATGACC-3′ and 5′-ATGCCACAGGATTCCATACCC-3′. Autoradiograms were performed by exposure of radioactive membranes to a phosphoimager screen for 1–24 h.
Statistical analysis
Statistical analyses were performed using SPSS 10·0 for Windows (SPSS Inc., Chicago, IL, USA). The results are presented as mean values and standard deviations. The statistical one-way ANOVA and t test were used for statistical evaluation.
Results
Food intake
In the HFD groups, food intake tended to be lower by about 15 % than the NFD group in the first 2-week period that feed was changed from the NFD to the HFD; after the rats had adapted to the HFD, the food intake of the HFD rats was almost the same as the NFD group during the experimental period. As a result, rats in the two HF-fed groups consumed significantly more energy than rats in the NFD groups. Additionally, no significant difference was found in the food intake between the HFD (30·40 (sd 4·4) g/d per rat) and HFD+CP (31·39 (sd 6·2) g/d per rat) groups.
Body and epididymal fat pad weights
As indicated in Fig. 1(A), the body weight of the HFD group was significantly increased compared with the NFD group after 6 months of feeding (P < 0·05) and remained significantly increased for the duration of the experiment. Addition of chickpeas to the HFD reduced the weight gain, such that the HFD+CP group had a statistically significantly lower body weight (P < 0·05) from 6 months to the end of the experiment compared with the HFD group (NFD 584 (sd 31·2); HFD 654 (sd 45·2); HFD+CP 562 (sd 60·3) at 8 months).

Fig. 1 Body-weight gain and epididymal fat pad weight of rats fed one of three diets for 8 months. (A) Body-weight changes throughout the experimental period for rats fed a normal-fat diet (NFD; □), a high-fat diet (HFD; ■) or the HFD plus chickpeas (HFD+CP; ![]() ) for 8 months. (B) Epididymal fat pad weight:total body weight ratio after feeding with the NFD, HFD or HFD+CP for 8 months. Values are means, with standard deviations represented by vertical bars. Mean value was significantly different from that of the HFD group: *P < 0·05, **P < 0·01.
) for 8 months. (B) Epididymal fat pad weight:total body weight ratio after feeding with the NFD, HFD or HFD+CP for 8 months. Values are means, with standard deviations represented by vertical bars. Mean value was significantly different from that of the HFD group: *P < 0·05, **P < 0·01.
The epididymal fat pad weight (an index of visceral adiposity in ratsReference Brand-Miller, Holt, Pawlak and McMillan22, Reference Yajima, Noguchi, Ikeshima, Shiraki, Kanaya, Tsuboyama-Kasaoka, Ezaki, Oikawa and Kondo23) v. total body weight of animals was significantly different among the three groups (Fig. 1(B)). It was higher in the HFD group (0·032 (sd 0·0042) g/g) than in both the NFD group (0·015 (sd 0·0064) g/g) and the HFD+CP group (0·023 (sd 0·0072) g/g; P < 0·05 compared with the HFD group).
Biochemical analysis for serum parameters
As indicated in Table 1, serum TAG, LDL-C, leptin and LDL-C:HDL-C levels were increased in the HFD rats, compared with those fed a standard diet. However, compared with the HFD group, chickpea treatment resulted in a 45 % decrease in the serum TAG, a 23 % decrease in LDL-C, a 35 % rise in HDL-C and a 30 % reduction in LDL-C:HDL-C. The serum leptin levels were also lower in the chickpea-fed group compared with the HFD group, but this did not reach significance. Unexpectedly, the HFD in the present experiment did not alter the serum level of total cholesterol.
Table 1 Biochemical analysis of serum parameters after the feeding period of 8 months (Mean values and standard deviations)
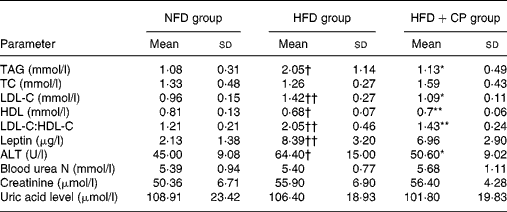
NFD, normal-fat diet; HFD, high-fat diet; HFD+CP, high-fat plus chickpea diet; TC, total cholesterol; LDL-C, LDL-cholesterol; HDL-C, HDL-cholesterol; ALT, alanine aminotransferase.
Mean value was significantly different from that of the HFD group: *P < 0·05, **P < 0·01.
Mean value was significantly different from that of the NFD group: †P < 0·05, ††P < 0·01.
On the other hand, the long-term HFD induced a slight increase of serum alanine aminotransferase levels and this tendency disappeared in rats treated with chickpeas, which suggested that chickpeas had some hepatic protective effect in this regard. The serum blood urea N, creatinine and uric acid level showed no difference among the three groups.
Lipoprotein lipase activities and triacylglycerol levels of tissues
Rats fed the HFD had higher TAG concentration in muscle and liver tissue, whereas the addition of chickpeas drastically lowered TAG (muscle by 39 % and liver by 23 %). The activities of LPL in epididymal fat and HTGL in liver increased by a 40 and 23 % respectively in the HFD rats compared with NFD rats; however, dietary chickpeas completely normalised the levels (Table 2).
Table 2 Lipoprotein lipase (LPL) activities as well as triacylglycerol levels in tissues after chickpea administration for 8 months (Mean values and standard deviations)
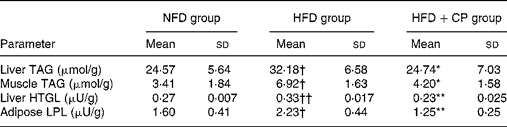
NFD, normal-fat diet; HFD, high-fat diet; HFD+CP, high-fat plus chickpea diet; HTGL, hepatic TAG lipase.
Mean value was significantly different from that of the HFD group: *P < 0·05, **P < 0·01.
Mean value was significantly different from that of the NFD group: †P < 0·05, ††P < 0·01.
Leptin and lipoprotein lipase mRNA expression
Northern blot hybridisation of epididymal adipose tissue total RNA with cDNA probes for leptin and LPL mRNA is shown in Fig. 2. Long-term high-fat feeding induced the higher leptin and LPL mRNA expression, as reported in previous studiesReference Zabala, Churruca, Fernandez-Quintela, Rodriguez, Macarulla, Martinez and Portillo24, Reference Takahashi and Ide25. However, chickpea-treated obese rats showed a markedly lower leptin and LPL mRNA content than untreated HFD groups.

Fig. 2 Representative Northern blots for mRNA of (A) leptin and (B) lipoprotein lipase (LPL) in epididymal adipose tissue of rats that had been fed a normal-fat diet (NFD), a high-fat diet (HFD) or the HFD plus chickpeas (HFD+CP) for 8 months. Total RNA (20 μg per lane) was analysed. The corresponding blot for the β-actin RNA is also shown. Values are means, with standard deviations represented by vertical bars. * Mean value was significantly different from that of the HFD group (P < 0·05).
Insulin tolerance test and oral glucose tolerance test
Basal plasma glucose levels showed no difference among the three groups after 7 months of diet supplementation. In response to insulin (ITT), plasma glucose levels at 60, 90 and 120 min in the HFD group were significantly higher than those in the NFD group (P < 0·05). However, the HFD+CP group had a hypoglycaemic response compared with the HFD group, and no statistical differences were found between the NFD+CP group and NFD group at any time point (Fig. 3(A)). Furthermore, during the 120 min following glucose administration (Figs. 3(B) and (C)), plasma glucose levels in the HFD group were significantly higher than those in the control group at 60, 90 and 120 min, indicating that the rats in the HFD group had developed impaired glucose tolerance. However, in this test the responses recorded in the NFD+CP group did not reach those of the NFD controls, and increased responsiveness in the insulin-releasing test was even evident at the early phase of the experiment though it did not reach statistical significance. Nevertheless, postprandial levels of plasma glucose (90 and 120 min) and insulin (120 min) in chickpea-treated rats were much lower than those in HFD-induced animals. Blood glucose was 6·68 (sd 1·02) mmol/l in the HFD+CP group compared with 8·94 (sd 1·08) mmol/l in the HFD group (P < 0·01) and plasma insulin was 0·18 (sd 0·041) nmol/l in the HFD+CP group compared with 0·23 (sd 0·03) nmol/l in the HFD group (P < 0·05) at 120 min. Therefore, the administration of chickpeas prevented postprandial hyperglycaemia and hyperinsulinaemia induced by a long-term HFD.

Fig. 3 Insulin resistance in rats fed a normal-fat diet (NFD; Δ), a high-fat diet (HFD; ▲) or the HFD plus chickpeas (HFD+CP; ■). (A) Insulin tolerance test (2 U/kg body weight; intraperitoneal) in rats fed the NFD, HFD or HFD+CP for 5 months. Changes in plasma glucose (B) and insulin (C) in rats during oral glucose tolerance tests (2 g/kg body weight) performed after 7 months on the different diets. Values are means, with standard deviations represented by vertical bars. Mean value of the HFD+CP group was significantly different from that of the HFD group: *P < 0·05, **P < 0·01.
Discussion
Rats fed a HFD over an extended time became obese, dyslipaemic and insulin resistant – findings consistent with previous reportsReference Tsutsumi, Inoue, Shima, Iwasaki, Kawamura and Murase26, Reference Kishino, Ito, Fujita and Kiuchi27. The present study focused on the possible beneficial effect of chickpeas to reverse visceral adiposity, hyperlipidaemia and impaired glucose tolerance in high-fat-fed rats. In the present study increases in the body weight and weight of epididymal adipose tissues were successfully prevented by chickpea supplementation to HFD-fed rats. Furthermore, effects of dietary chickpeas on adipose tissue are not simply a reflection of decreased body weight because adipose decreases exceeded body-weight changes (Fig. 1). Chickpea treatment also resulted in the significant decrease of a number of pro-atherogenic factors, such as TAG, LDL-C and LDL-C:HDL-C. In addition, as adipocyte mRNA marker is closely related to adipose cell hypertrophyReference Raclot, Groscolas, Langin and Ferre28, Reference Ailhaud, Grimaldi and Negrel29, the lower expression of both LPL (the early differentiation marker) and leptin (the late differentiation markerReference Ailhaud, Grimaldi and Negrel29) in our experiments suggests a modulation of the adipocyte phenotye in the chickpea group.
Of the various body fat depots, the amount of visceral fat is positively correlated to the insulin sensitivity in animal models and man; insulin action is markedly impaired in individuals with visceral obesityReference Axen, Dikeakos and Sclafani30, Reference Carey, Jenkins, Campbell, Freund and Chisholm31. Meanwhile, ectopic lipid deposition in muscle and liver is also postulated to mediate insulin resistanceReference McGarry32, with increased intramyocellular lipids contributing to the development of type 2 diabetesReference Kelley, Goodpaster and Storlien33, and hepatic TAG content strongly associating with hepatic insulin resistanceReference Utzschneider and Kahn34. So, the hypolipidaemic and insulin-sensitising action of chickpeas in high-fat-fed rats may partly be due to the reduction of visceral and ectopic fat depots.
Similar favourable modifications in lipid profiles by chickpea feeding have been observed in several studies. Zulet et al. Reference Zulet, Macarulla, Portillo, Noel-Suberville, Higueret and Martinez18 have showed that chickpea feeding significantly decreased serum total LDL-C and TAG levels and increased hepatic glucokinase activity in rats fed on a hypercholesterolaemic diet containing coconut oil and cholesterolReference Zulet, Macarulla, Portillo, Noel-Suberville, Higueret and Martinez18, Reference Zulet and Martinez35. Wang & McIntoshReference Wang and McIntosh6 have found a cholesterol-lowering ability of chickpeas, both processed by boiling and extrusion. Others have also reported that dietary chickpeas successfully decreased TAG and cholesterol concentrations in the heart and liver tissue of ratsReference Habib and Ahsan17, Reference Muzaffar, Bakhsh, Khan and Khan36.
Moreover, the present results are in agreement with previous work demonstrating that high-fat feeding increases LPL and HTGL activity in white adipose tissue and liverReference Ait Yahia, Madani, Prost, Bouchenak and Belleville37. However, LPL and HTGL activities were found to be significantly reduced in HFD+CP rats; this might explain the lowered lipids in adipose tissue and liver in the HFD+CP group compared with the HFD group. In obese subjects, lowering of fat accumulation and reducing the circulating TAG levels are necessary in order to improve fat metabolism. Recently Naaz et al. Reference Naaz, Yellayi, Zakroczymski, Bunick, Doerge, Lubahn, Helferich and Cooke38 have shown that the soya isoflavone genistein decreases adipose deposition by decreasing LPL mRNA in female miceReference Naaz, Yellayi, Zakroczymski, Bunick, Doerge, Lubahn, Helferich and Cooke38. Lee also found that soya protein and genistein improved blood glucose, antioxidant enzyme activities, and lipid profile in streptozotocin-induced diabetic ratsReference Lee39. Therefore it is possible that some components of chickpea isoflavone may be responsible for the effects seen in the present study. In addition, the lipid fraction from chickpeas is high in unsaturated fatty acids, primarily linoleic and oleic acids, which may have a synergistic effect of modifying body composition and plasma lipids.
Despite no statistically different basal plasma glucose levels among NFD, HFD and HFD+CP groups, the present results still indicate that chickpea-supplemented rats are protected from HFD-induced impairment of glucose tolerance and insulin resistance (Fig. 2). We hypothesise that the hypoglycaemic effect of chickpeas in the ITT and the oral glucose tolerance test were secondary to the metabolic changes associated with reduced adiposity. To the best of our knowledge, the present study is the first to demonstrate that dietary chickpeas improved insulin resistance (in the ITT) and reversed impaired glucose intolerance (in the oral glucose tolerance test) in long-term high-fat-fed animals. In healthy human subjects, both Johnson et al. Reference Johnson, Thomas and Hall16 and Nestel et al. Reference Nestel, Cehun and Chronopoulos15 found some hypoglycaemic effects after a single chickpea-based breakfast. However, no significant effects were obtained after a long-term (6 weeks) treatment according to Nestel et al. Reference Nestel, Cehun and Chronopoulos15; the lack of change could be due to the selection of normal subjects in this studyReference Nestel, Cehun and Chronopoulos15, Reference Johnson, Thomas and Hall16. Further research is necessary to demonstrate if chickpeas improve glucose disposal in insulin-resistant and diabetic subjects.
In conclusion, the results from the present study show that the chickpea has potential to assist with the prevention of excess fat accumulation induced by a chronic HFD and subsequently to prevent hyperlipidaemia as well as diabetes. The present findings provide a rational basis for the consumption of chickpeas as a functional food ingredient, which may be beneficial for correcting obesity-related diseases.
Acknowledgements
The present study was supported by the Shanghai-Unilever Research and Development Fund (2004) and a grant from the cultivation programme of Shanghai excellence young medical talent fund (2004). The present study was also partially supported by Jumpsun Bio-medicine (Shanghai) Co. Ltd. Y. Y. and L. Z. contributed equally to the present study.





