



Introduction
The Brazilian Cerrado has stood out as one of the most important regions in the farming sector, notwithstanding, soil fertility does not follow the same merit. Agricultural crops are highly dependent on and responsive to mineral fertilization since it improves forage accumulation (FA), nutritive value and animal weight gain. The integration between crops and livestock may minimize the input dependency, and produce sustainable food (Sekaran et al., Reference Sekaran, Lai, Ussiri, Kumar and Clay2021) while promoting ecosystem services (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017). In this context, integrated crop–livestock systems (ICLS) have followed a trend of improving productivity as indicated by shifts in soil biological, physical and chemical properties (Carvalho et al., Reference Carvalho, Neto, Barro and Nunes2018; Deiss et al., Reference Deiss, Kleina, Moraes, Franzluebbers, Motta, Dieckow, Sandini, Anghinoni and Carvalho2019).
Other benefits derived from the adoption of ICLS besides those already mentioned include lower greenhouse gases emissions (Khan et al., Reference Khan, Abbas AFarooque, Abbas and Wang2022), more efficient use of land and machinery, weed suppression (MacLaren et al., Reference MacLaren, Storkey, Strauss, Swanespoel and Schmutz-Dehnen2018), reduction in the incidence of diseases in agricultural crops (Wang et al., Reference Wang, Pan, Yang, Burdon, Friberg, Sui and Zhan2021) and improvements in economic return to the producer (Vinholis et al., Reference Vinholis, Filho, Shimata, Oliveira and Pedroso2021), especially in tropical regions such as the Brazilian Savannah (i.e. Cerrado) (Pariz et al., Reference Pariz, Costa, Crusciol, Castilhos, Meirelles, Roça, Pinheiro, Kuwahara, Martello, Cavasano, Yasuoka, Sarto, Melo and Franzluebbers2017).
Part of nutrients can be exported by crops and animals in ICLS. For example, soybean crops exported 66.0, 4.6 and 16.1 kg/Mg of grains produced of N, P and K, respectively (Francisco et al., Reference Francisco, Câmara and Segatelli2007), in contrast, 5–30% were exported by grazing animals as animal product (Rotz et al., Reference Rotz, Taube, Russelle, Oenema, Sanderson and Wachendorf2005). In general, 70–90% of the nutrients ingested by ruminants return to the pasture via urine and dung (Vendramini et al., Reference Vendramini, Dubeux and Silveira2014), nutrients not exported by animals can be reused by the subsequent agricultural crop.
Nutrient cycling occurs intensely in ICLS and the dynamics of this cycling is intensified by the presence of grazing animals (Carvalho et al., Reference Carvalho, Anghinoni, Moraes, Souza, Sulc, Lang, Flores, Lopes, Silva, Conte, Wesp, Levien, Fontaneli and Bayer2010). Therefore, the animals catalyse nutrient cycling (Moraes et al., Reference Moraes, Carvalho, Anghinoni, Lustosa, Costa and Kunrath2014), because they partially decouple N and C when they consume forage (Lemaire et al., Reference Lemaire, Gastal, Franzluebbers and Chabbi2015). This nutrient cycling promoted by animals can be used to reduce fertilizer costs (Kronberg, Reference Kronberg2021) and develop more sustainable production systems.
Among nutrients, N is often the one that most limits plant growth (Hogberg et al., Reference Hogberg, Nasholm, Franklin and Hogberg2017). N increases leaf appearance and expansion rates (Costa et al., Reference Costa, Moraes, Monteiro, Motta, Oliveira and Rodrigues2013), the photosynthetic potential of leaves and canopy (Yasuoka et al., Reference Yasuoka, Pedreira, Silva, Alonso, Silva and Gomes2018), great animal productivity (Gomes da Silva et al., Reference Gomes da Silva, Dubeux, de Mello, da Cunha, dos Santos, Apolinário and de Freitas2021) and accumulation rate of leaves and stem (Silva et al., Reference Silva, Pedreira, Sollenberger, Silva, Yasuoka and Almeida2016a). Bernardon et al. (Reference Bernardon, Assmann, Soares, Franzluebbers, Maccari and Bortolli2021) consider N application on grazing phase in ICLS essential to ensure the appropriate growth of forage, also improving forage nutritive value (Silva et al., Reference Silva, Pedreira, Sollenberger, Silva, Yasuoka and Almeida2016b). The increase in production performance of tropical forage plants in response to N application is widely documented (Gurgel et al., Reference Gurgel, Difante, Montagner, Araújo, Dias, Santana, Rodrigues and Pereira2020; Oliveira et al., Reference Oliveira, Correa, Cunha, Rêgo, Faturi, Silva and Domingues2020). Accordingly, the application of N can improve the production performance of forage plants and ensure the long-term sustainability of pasture (Borges et al., Reference Borges, Silveira, Cardoso, Moline, Coutinho Neto, Lucas, Muraoka and Coutinho2017).
Fertilization in ICLS is still based on conventional cultivation, disregarding the nutrient flow that occurs in the system. However, nutrients can be better utilized when fertilizers are applied at more appropriate times. It is in this context that the system fertilization presented by Farias et al. (Reference Farias, Dubeux, Savian, Duarte, Martins, Tiecher, Alves, de Faccio Carvalho and Bremm2020) emerges, differing from the traditional forms of fertilization in ICLS, which seek to maximize only the production of the agricultural crop. This strategy of fertilization suggests the application of nutrients in the pasture phase because there is less nutrient extraction. Thus, the intense cycling of nutrients promoted by the animals will cause the residual effect of fertilization to be used by the subsequent crop. In this way, it is possible to avoid losses and achieve maximum efficiency in the use of nutrients, maximizing the total production of the system.
The reallocation of phosphorous and potassium from the crop to the pasture phase, and nitrogen fertilization in the pasture, also was investigated in the same area of this research by Pires et al. (Reference Pires, Denardin, Silva, Freitas, Gonçalves, Camargo, Bremm, Carvalho and Souza2022). The authors related the higher soybean yields to improvements in soil biochemical and biological properties. The use of SF increased the efficiency of carbon and phosphorus use due to increased activities of β-glucosidase and acid phosphatase. Nonetheless, data showing the effect of these strategies in the pasture phase are still needed to analyse the global efficiency of this new concept of fertilization.
For this reason, the aim of this research was, therefore, to evaluate how the fertilization strategy (P and K application in soybean or pasture) associated with or without nitrogen fertilization in the pasture phase affects the accumulation and chemical composition of forage and its implications on animal productivity in ICLS.
Material and methods
Site, climate and soil description
The research was conducted in the Guatita Farm (16°33'54” S, 54°41'08” W, 227 m a.s.l.), located in Rondonópolis, MT-Brazil from March 2019 to July 2020. The climate is classified as Aw according to Köppen's classification (Alvares et al., Reference Alvares, Stape, Sentelhas, Gonçalves and Sparovek2013). The climatic data of the experimental period are presented in Fig. 1. The soil of the experimental area is a Latossolo Vermelho distrófico, which is equivalent to an Oxisol according to the soil survey staff. The chemical characteristics of the soil for the 0–20 cm layer before the experiment (February 2019) were: pH (CaCl2) = 5.5; organic matter = 15.1 g/dm3;Ca = 2.5 cmolc/dm3; Mg = 1.3 cmolc/dm3; Al = 0.0 cmolc/dm3, cation exchange capacity = 6.8 cmolc/dm3; K = 69.2 cmolc/dm3; P = 30.8 cmolc/dm3 and base saturation = 59.9%. The proportions of sand, silt and clay were 505, 75 and 420 g/kg, respectively, classifying the soil as of clay texture, and bulk density was 1.4 kg/dm3, which is considered moderate.

Figure 1. Total rainfall (mm) and maximum and minimum temperatures (°C) during the experimental period in 2019 and 2020.
Experimental design and treatments
The experiment was conducted in an ICLS and treatments were distributed in randomized blocks with three replications and four fertilization strategies, totaling 12 experimental units (paddocks) of 1.9 ha each (total of 22.8 ha).
1. Conventional fertilization with P and K on grain crop without nitrogen in the pasture phase (CF–N); 2. Conventional fertilization with P and K on grain crop with nitrogen in the pasture phase (CF + N); 3. Pasture fertilization with P and K in the pasture phase (SF–N); 4. System fertilization with N, P and K in the pasture phase (SF + N). In the CF + N phosphorus and potassium were applied to soybean sowing and nitrogen at the pasture establishment. Phosphate and potassium fertilization was calculated for an estimated yield of 4.0 Mg/ha of soybean grains. For this, total quantities of 31 kg P/ha and 75 kg K/ha were applied using triple superphosphate and potassium chloride, respectively. The cultural practices of soybean cultivation followed the technical recommendations for the region. In the treatment with N application, the dose of 100 kg N/ha was applied using urea and a nitric and ammoniacal source in the first and second experimental year, respectively. N application was carried out when the plants reached four fully expanded leaves, which occurred approximately 15 days after pasture sowing.
The soybean [Glycine max (L.) Merr.] cultivar Brasmax Ultra IPRO was sown at a spacing of 50 cm between rows in October 2018, in a no-tillage system directly in the straw. The experimental period (year 1) began by sowing the pasture (Urochloa brizantha (Hochst. ex A. Rich.) R. D. Webster [syn. Brachiaria brizantha (Hochst. ex A. Rich.) Stapf] cv. BRS Piatã on 18 March 2019, as soon as soybean was harvested. Soybean was again sown on 12 October 2019, using the same cultivar and the same planting system as the previous year and harvested on 24 February 2020, when Piatã palisade grass was sown again (year 2). The pasture formed was evaluated until July 2020, when the climatic conditions stopped the growth of the forage and, for this reason, the experiment was ended. In both years, Piatã palisade grass was sown with a small-grain seeder using 6 kg/ha of pure and viable seeds with a spacing of 17 cm between rows.
Livestock management and responses
Nellore male cattle with an initial average weight of 246 ± 8.1 kg and 260 ± 6.2 kg in years 1 and 2, respectively, were managed under continuous stocking with variable stocking rate, using ‘put and take’ technique with three tester animals fixed in each paddock. These animals remained in the area from April 13 to August 23, 2019, and from March 21 to July 17, 2020, totaling 132 and 118 days of grazing in years 1 and 2, respectively. The shorter grazing period in year 2 was due to lower rainfall (Fig. 1). The animals were placed in the area when the canopy height was ~35 cm. Grazing management aimed to maintain the canopy height around 32 cm, close to that used by Euclides et al. (Reference Euclides, Montagner, Barbosa, do Valle and Nantes2016), which was 30 cm. The amplitude allowed ranged from ~24 to 40 cm (Table 1), within the range from 15 to 45 cm recommended by Nantes et al. (Reference Nantes, Euclides, Montagner, Lempp, Barbosa and Gois2013) for Piata palisade grass. The biggest concern was not to let the average height exceed 40 cm. A variable number of grazer animals similar in breed and weight to testers were added or removed to keep the height within the pre-established limits. All animals had free access to water and mineral salt throughout the experimental period.
Table 1. Averages and standard deviation of pasture height for each treatment in 2019 and 2020

CF – N, crop fertilization without N in the pasture; CF + N, crop fertilization with N in the pasture; SF – N, system fertilization without N in the pasture; SF + N, system fertilization with N in the pasture; ns, not significant.
Tester and grazer animals were weighed monthly to adjust the stocking rate. The weighing procedures were after feed and water fasting of 16 h. The average daily gain (ADG) was calculated by the difference between the final and initial weight of the tester animals divided by the number of days of evaluation. Gain per area (GPA) was determined by the difference between the sum of the values resulting from the final weight and the initial weight of the tester and put-and-take animals.
The stocking rate (SR) was calculated as the sum of the body weight (BW) of tester and put-and-take animals expressed per area unit (ha) and per animal unit (AU), representing 450 kg BW. Weight gain per area was calculated as the ADG multiplied by SR and the number of days of the evaluated period.
Supplementation was provided once a day in the morning to ensure the maintenance of the three tester animals until the end of the evaluation period (Table 2), due to the drop in nutritional value and forage accumulation during the lower rainfall season (Fig. 1). Supplementation was provided when the forage had CP below 7% in both years (Table 2). In the first year, supplementation was 0.3% BW after 86 days of grazing. In the second year, supplementation began after 72 days of grazing, initially at 0.5% BW and gradually increasing to 0.8% BW until the end of the grazing period. Supplementation consisted of 85% supplement plus 15% of white salt for regulating the food intake. The higher level of supplementation in year 2 was due to the longer period without rainfall.
Table 2. Composition of the supplement used during the experiment in both experimental years

DM, dry matter.
1 Guaranteed levels: 115 g/kg Ca; 20 g/kg P; 30 g/kg Mg; 115 g/kg Na; 40 g/kg S; 1000 mg/kg Cu; 3000 mg/kg Zn; 2000 mg/kg Mn; 75 mg/kg I; 18 mg/kg Se; 60 mg/kg Co; 30 mg/kg Cr; 220000 IU/kg Vit A; 27500 IU/kg Vit D3; 1000 IU/kg Vit E; 2500 mg/g monensin; 1500 mg/kg virginiamycin.
Forage accumulation
Three sites representing the average condition of the canopy were selected in each experimental unit for estimating FA. An exclusion cage with dimensions of 1.0 × 1.0 × 1.2 m width, length and height, respectively, was placed at each of these points. Forage mass (FM) was determined on the same date when cages were installed, using the double-sampling technique and direct measurements were obtained by harvesting six 0.25/m2 quadrats per paddock at ground level. Forage samples were oven-dried at 60 °C for 72 h to a constant weight. On the last day of each accumulation cycle, the FM within each exclusion cage was estimated using the same procedure previously reported. After sampling, cages were placed on sites representing the average pasture structure and relocated every eight up to 28 days depending on the growth rate of the plants, shorter when climatic condition was less favourable and longer when it was more favourable. Forage accumulation was calculated as the FM inside the cage minus the FM in the paddock at the time the cages were placed. Total forage accumulation (total FA) was calculated as the sum across all FA cycles. Forage accumulation divided by the growth period, resulted in the forage accumulation rate (FAR). When forage accumulation had negative values, it was considered zero.
Morphological and chemical composition of forage
Morphological composition was evaluated monthly by collecting samples at three representative points forage mass average of the paddock. At these points, all the forage contained within 0.25/m2 frames was collected at ground level, separated into leaf (leaf blade), stem (stem + leaf sheath) and dead material. After separation, all components were dried separately in a forced air circulation oven at 60°C to a constant weight. The dry weight of each component was used to determine the relative proportion of leaf, stem and dead material in FM.
For the chemical composition of forage, hand-plucked samples were collected monthly. The samples were dried in a forced air circulation oven at 60°C to constant weight. After drying, samples were ground to pass a 1-mm screen using a Wiley Mill. Total N concentration and crude protein content (CP) were calculated as the total N × 6.25 using the Kjeldahl method. Neutral detergent fibre (NDF), acid detergent fibre (ADF) and lignin contents were determined by following the methodology proposed by Silva and Queiroz (Reference Silva and Queiroz2006).
Data analysis
Data were submitted to analyses of variance using the mixed procedure of the statistical package SAS (version 9.4, SAS Institute), including the effects of the fertilization strategy. Year and block were considered random effects. The covariance structure was chosen using Akaike's information criterion (AIC). Linear prediction and quantile–quantile plots of the residuals were used to check the normality and homogeneity of variance of the residuals. Treatment means were estimated using the ‘LSMEANS’ and compared by the probability of difference (PDIFF), using Student's t-test (P < 0.05). Monthly averages were calculated for chemical composition and morphological composition and the values obtained were used in the statistical analysis.
Principal component analysis (PCA) was used to establish attributes with discriminatory power and potential indicators of the effect of FA, chemical composition, animal performance and gain per area under fertilization strategies in ICLS. The relationships of the variables explained by a limited number of new variables, eliminating the effect of collinearity, were calculated from the correlation matrix between variables. The original values were normalized to mean equal to 0 and variance equal to 1. The criterion adopted for choosing the number of components was to select those that had eigenvalues above 1.00 and were able to synthesize an accumulated variance above 70%.
Results
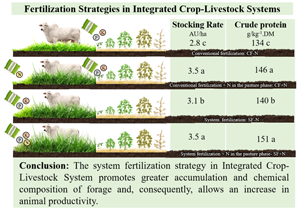
Total FA and FAR were affected by the fertilization strategy (P = 0.0010 and P = 0.0012, respectively) (Table 3). System fertilization treatment showed the highest values compared to the others. Total FA in SF + N was 25% higher compared to CF–N, a raise of 5210 kg DM ha/year. The values in CF + N, CF–N and SF–N were similar.
Table 3. Forage accumulation and forage accumulation rate of Piatã palisade grass in an integrated crop-livestock system as affected by crop fertilization or system fertilization associated or not with nitrogen over two years in Brazilian Cerrado

DM, dry matter; CF–N, crop fertilization without N in the pasture; CF + N, crop fertilization with N in the pasture; SF–N, system fertilization without N in the pasture; SF + N, system fertilization with N in the pasture; SEM, standard error of the mean.
Means followed by a common letter in a column do not differ by t test (P > 0.05).
Contents of CP, NDF and ADF on stem and dead material proportions were affected by the fertilization strategy (P = 0.0002, P = 0.0302, P = 0.0456, P = 0.0180 and P = 0.0004, respectively). The CP was higher in treatments with nitrogen application when compared to those that did not receive N. The NDF was higher in pasture fertilized without N followed by crop fertilized with N and crop fertilized without N. The ADF and the proportion of stem were higher in CF – N and SF – N and lower in CF + N and SF. The proportion of dead material was higher in SF and lower in CF – N. The CF + N and SF–N showed intermediate values. Neutral detergent fibre was higher in SF – N and lower in CF–N and CF + N (Table 4). Lignin content and the proportion of leaves were not affected by treatments (P = 0.3585 and P = 0.1252, respectively), with mean values of 29.2 g/kg DM and 29.3%, respectively.
Table 4. Crude protein (CP), neutral detergent fibre (NDF), acid detergent fibre (ADF), and stem and dead material proportions in an integrated crop–livestock system as affected by crop fertilization or system fertilization associated or not with nitrogen over two years in Brazilian Cerrado

DM, dry matter; CF – N, crop fertilization without N in the pasture; CF + N, crop fertilization with N in the pasture; SF – N, system fertilization without N in the pasture; SF + N, system fertilization with N in the pasture; SEM, standard error of the mean.
Means followed by a common letter in a column do not differ by t test (P > 0.05).
The ADG, SR and GPA were affected by the fertilization strategy (P = 0.0317, P = 0.0308, P = 0.0001 and P < 0.0001). The ADG was lower in SF – N compared to the CF + N, CF – N and SF treatments, which were similar to each other. The SR and GPA were higher in treatments with N (CF + N and SF), followed by SF – N. The CF – N had the lowest values of SR and GPA (Table 5).
Table 5. Average daily gain (ADG), stocking rate (SR), and gain per area (GPA) in an integrated crop–livestock system as affected by crop fertilization or system fertilization associated or not with nitrogen over two years in Brazilian Cerrado

LW, live weight; AU, animal unity; BW, body weight; CF–N, crop fertilization without N in the pasture; CF + N, crop fertilization with N in the pasture; SF–N, system fertilization without N in the pasture; SF + N, system fertilization with N in the pasture; SEM, standard error of the mean.
Means followed by a common letter in a column do not differ by t test (P > 0.05).
1 AU: animal unity of 450 kg BW.
The PCA explained 77% (53% in PC1 and 24% in PC2) of the total variance, with eigenvalues above 1.0. The PC1 was defined by the variables correlated with forage accumulation and animal productivity (Fig. 2A) according to the following order of importance contribution: CP (−0.93) > SR (−0.92) > GPA (−0.86) > proportion of stem (0.84) >FA (−0.82) > proportion of dead material (−0.81) (Fig. 2B). In contrast, NDF (−0.89) > proportion of leaves (0.73) > ADG (0.72) characterized PC2. The fertilization strategies with N application in the pasture (CF + N and SF + N) were associated with higher values of forage accumulation, SR, GPA, CP and proportion of dead material, which are positively correlated variables. These variables are inversely correlated with the proportion of stem, which was associated with treatments without N (CF–N and SF–N) (Fig. 2A).

Figure 2. Principal component analysis (A) and contribution of variables (B) related to the chemical and morphological composition of forage, forage accumulation and animal productivity in Piatã palisade grass pastures subjected to different fertilization strategies in integrated agricultural production systems. CF, crop fertilization without N in the pasture; CFN, crop fertilization with N in the pasture; PF, system fertilization without N in the pasture; SF, system fertilization with N in the pasture; NDF, neutral detergent fibre.
Discussion
Forage accumulation rate ranged from 126.5 to 169.3 kg DM/ha.day (Table 3). These values can be considered higher than previous studies because the experiment began at the end of the rainy season and extended over the dry period. Euclides et al. (Reference Euclides, Montagner, Barbosa, do Valle and Nantes2016) evaluated Piatã palisade grass fertilized with 90 kg N/ha kept under constant height of 30 cm during the rainy season and reported a FAR of 64.1 kg DM/ha.day. Machado et al. (Reference Machado, Sales, Reis, Mesquita, Carvalho, Monção, Gomes, Queiroz and Lima2017) found a linear increased in FAR of Brachiaria decumbens Stapf (syn. Urochloa decumbens (Stapf) Webster cv. Basilisk) in response to the applied N doses, reaching from 117, 137, 167, 171 to 216 kg DM/ha.day when the N doses were 0, 100, 200, 300 and 400 kg/ha, respectively.
The higher forage accumulation in the treatments CF + N and SF + N (Fig. 2A) was due to nitrogen fertilization since the grazing management was the same between treatments. A similar argument was used by Delevatti et al. (Reference Delevatti, Cardoso, Barbero, Leite, Romanzini, Ruggieri and Reis2019), who evaluated the effect of four N doses (0, 90, 180 and 270 kg N/ha) on Marandu palisade grass. The authors stated that nitrogen fertilization was responsible for the increase in FA since the pasture management height was the same for all treatments (25 cm kept constant). Increase in the production performance of Piatã palisade grass in response to nitrogen fertilization was expected since, among nutrients, N is the main determinant of dry matter production, being responsible for maintaining the productivity and sustainability of the pasture ecosystem (Avelino et al., Reference Avelino, Faria, Penso, Lima, Rodrigues, Abreu, Cabral and Peixoto2019). After being assimilated, N associates with carbon chains, increasing cellular constituents, improving regrowth vigour and increasing forage production (Galindo et al., Reference Galindo, Buzetti, Teixeira Filho, Dupas and Ludkiewicz2017). Moreover, there is a direct relationship between the accumulation of dry matter by the plant and the increase in atmospheric CO2 fixation (Taiz et al., Reference Taiz, Zeiger, Moller and Murphy2017).
Nitrogen impacts physiological responses, tillering dynamics and canopy structure, resulting in increased forage accumulation. In a study that evaluated Mulatto II brachiaria grass under two N doses (50 and 250 kg/ha/year), the application of the highest N dose increased tiller appearance rate, tiller population density (Silva et al., Reference Silva, Silva, Yasuok, Sollenberger and Pedreira2020), leaf and canopy photosynthesis rates, leaf area index, canopy light interception (Yasuoka et al., Reference Yasuoka, Pedreira, Silva, Alonso, Silva and Gomes2018) and leaf and stem accumulation rates (Silva et al., Reference Silva, Pedreira, Sollenberger, Silva, Yasuoka and Almeida2016a). The results of the present study together with those already consolidated in the literature confirm the statement of Dupas et al. (Reference Dupas, Buzetti, Rabêlo, Sarto, Cheng, Teixeira Filho, Galindo, Dinalli and Gazola2016) and Avelino et al. (Reference Avelino, Faria, Penso, Lima, Rodrigues, Abreu, Cabral and Peixoto2019), that N is the nutrient that most affects forage accumulation. According to Bernardon et al. (Reference Bernardon, Assmann, Soares, Franzluebbers, Maccari and Bortolli2021), for ICLS to be successful, it is essential that N fertilization is carried out in the pasture phase, which is also confirmed by the results of the present study (Fig. 2A).
The application of N increased CP from 134 to 146 g/kg DM in conventional fertilization and from 140 to 151 g/kg DM in system fertilization (Table 4). The PCA analysis (Fig. 2A) demonstrates the impact of N fertilization on the increase in forage CP even in a period of low water availability (Fig. 1).
Several studies have shown that N fertilization reduces forage NDF Cecato et al. (Reference Cecato, Pereira and Jobim2004), Benett et al. (Reference Benett, Buzetti, Silva, Bergamaschine and Fabricio2008), Dupas et al. (Reference Dupas, Buzetti, Sarto, Hernandez and Bergamaschine2010), Dupas et al. (Reference Dupas, Buzetti, Rabêlo, Sarto, Cheng, Teixeira Filho, Galindo, Dinalli and Gazola2016) and Delevatti et al. (Reference Delevatti, Cardoso, Barbero, Leite, Romanzini, Ruggieri and Reis2019). In the present study, reduction in NDF caused by N application was not evident since there were no differences in NDF between the treatments SF + N and SF – N and between CF + N and CF – N (Table 4). Differences between these results and those previously mentioned may be related to the N dose applied. Dupas et al. (Reference Dupas, Buzetti, Sarto, Hernandez and Bergamaschine2010), Dupas et al. (Reference Dupas, Buzetti, Rabêlo, Sarto, Cheng, Teixeira Filho, Galindo, Dinalli and Gazola2016) and Delevatti et al. (Reference Delevatti, Cardoso, Barbero, Leite, Romanzini, Ruggieri and Reis2019) evaluated N doses ranging from 0 to 200, 700 and 270 kg/ha, respectively. The contrast between 0 and 100 kg N/ha may not have been sufficient to detect differences in NDF between treatments with and without N. Moreira et al. (Reference Moreira, Martuscllo, Fonseca, Mistura, Morais and Júnior2009) evaluated the effect of nitrogen on the levels of NDF of Brachiaria decumbens cv. Basilisk under nitrogen fertilization (75, 150, 225 and 300 kg/ha.year of N), in continuous stocking with variable stocking rate in two agricultural years consecutive and also did not find significant differences regarding nitrogen contents, only for months of evaluation. The authors attributed the limitation to environmental factors, mainly rainfall.
The highest proportion of dead material was associated with treatments with N (Fig. 2A). The increase in leaf and tiller renewal in treatments with N may be responsible for the higher proportion of dead material. Braz et al. (Reference Braz, Fonseca, Freitas, Martuscello, Santos, Santos and Pereira2011) found a negative linear correlation between N dose and leaf lifespan in Tanzania grass (Megathyrsus maximus (Jacq.) B. K. Simon and S. W. L. Jacobs) fertilized with 0, 80, 160 and 320 kg N/ha. Silva et al. (Reference Silva, Silva, Yasuok, Sollenberger and Pedreira2020), when studying the effects of two N doses (50 and 250 kg/ha) on the tillering dynamics of Mulatto II grass, observed that the highest N dose reduced tiller survival rate. In this case, the application of 100 kg/ha of N may have intensified the senescence rate of leaves and tillers, increasing the proportion of dead material in the forage. Also, the higher SR in these treatments might have increased the lodging of the grass, which resulted in higher plant damage and dead material proportion. As the proportion of leaves was similar between treatments, the higher proportions of dead material in SF + N and CF + N consequently resulted in a lower proportion of stem for these treatments (Table 4). There was a negative correlation between the proportions of stem and dead material, as shown in PCA analysis (Fig. 2A).
Regarding animal performance, the lower ADG in SF – N may have occurred due to NDF that was 2.6, 1.9 and 1.5% higher in this treatment compared to CF + N, CF – N and SF + N, respectively (Table 4). Although the values were similar, NDF was greater than 60% only in SF – N, a value from which it begins to limit the consumption of forage (Van Soest, Reference Van Soest1965). The high content of NDF reduces consumption because the digestion of the fibrous fraction of the plant is slow, causing the food to remain in the digestive tract for longer. Therefore, its content in forage is inversely related to the voluntary consumption of the animals. Negative correlation between NDF and ADG was found in the present study by PCA (Fig. 2A).
Although ADG was lower in SF – N, SR and GPA were higher in this treatment compared to CF – N (Table 5 and Fig. 2A), showing that the change in the time of application of P and K affected SR and GPA more than the individual performance of the animals. In a parallel study to the current one, Pires et al. (Reference Pires, Denardin, Silva, Freitas, Gonçalves, Camargo, Bremm, Carvalho and Souza2022) registered similar soybean yield in the first two years evaluated when P and K were applied in the pasture phase instead in the crop phase. The same authors also found soybean yield 16% higher when P, K and N were applied in the pasture compared to when P and K were applied in the crop phase without any N fertilization. These results demonstrate that fertilization in the pasture phase in ICLS (SF – N and SF) can improve animal productivity without reduction in soybean yield. This result is in line with the objective raised by Farias et al. (Reference Farias, Dubeux, Savian, Duarte, Martins, Tiecher, Alves, de Faccio Carvalho and Bremm2020) who postulated that the increase in food production must be associated with better efficiency in the use of nutrients.
The higher SR values in the two treatments containing N (CF + N and SF + N) compared to treatments without N (SF – N and CF – N) (Fig. 2A and Table 4) show that, in ICLS, the application of N in the pasture phase positively affects SR. Delevatti et al. (Reference Delevatti, Cardoso, Barbero, Leite, Romanzini, Ruggieri and Reis2019) observed a linear increase in SR from 3.4 to 6.6 AU/ha when evaluating N doses ranging from 0 to 270 kg/ha in Marandu palisade grass, demonstrating the major impact of N on the increase in SR. The authors state that the increase in forage accumulation in response to N fertilization was responsible for the increase in SR. The high correlation between forage accumulation and SR (Fig. 2A) shows that it occurred in the present study.
The higher SR in SF – N compared to CF – N (Table 5) demonstrates that when there is no application of N, changing the time of application of P and K from the crop phase to the pasture phase impacts the SR. Phosphorus is fundamental for the establishment of tropical pastures – which is the case of the present study in which the pasture was planted after the soybean crop – since this nutrient stimulates the growth and development of roots (Martins et al., Reference Martins, Monteiro and Pedreira2014). Potassium is the nutrient most required by plants after N and acts on stomatal opening and cell elongation (Hawkesford et al., Reference Hawkesford, Horst, Kichey, Lambers, Schjoerring, Møller, White and Marschner2012). Matos and Monteiro (Reference Matos and Monteiro1998) in a greenhouse study recorded an increase in forage accumulation in Marandu palisade grass as potassium rates increased. Probably, the application of P and K in the pasture phase may have led to greater availability of these nutrients to forage plants at the time of pasture establishment, resulting in greater forage accumulation (Fig. 2A) and, consequently, higher SR in SF–Ncompared to CF–N (Table 5).
The similar values of ADG between the treatments and the same response pattern of SR found in GPA, that is, higher values in CF + N and SF + N, followed by SF – N and CF – N with the lowest value (Table 5) show that GPA was determined by SR and not by ADG. The results of the present study reveal that both the application of N and the application of P and K in the pasture phase positively affect animal productivity. However, a better comprehension of the impact of these fertilization strategies would be possible if morphogenic, structural and physiological responses of plants and ingestive behaviour of animals were evaluated. Therefore, future studies evaluating these variables (e.g. leaf area index, light interception, leaf elongation rate, tiller population density, leaf photosynthesis rate, biting rate, etc.) may help a deep understanding of how the system fertilization affects the pasture phase in ICLS.
The CF + N had higher SR compared to the treatments CF – N and SF – N, although multivariate analysis showed that forage accumulation and forage accumulation rate were similar among these three treatments (Table 5). Nitrogen fertilization in CF + N was expected to result in higher forage accumulation compared to treatments without N (CF – N and SF – N). The PCA demonstrates that this result probably occurred since the CF + N treatment was more associated with forage accumulation than the treatments without N (Fig. 2A). Stocking rate is dependent on the amount of biomass produced and SR was higher in CF + N (Table 5). Nevertheless, the application of N in the pasture phase has higher impact to SR than ADG.
The mean values of ADG and SR were 0.779 kg LW/day and 3.2 AU/ha, respectively (Table 5). Euclides et al. (Reference Euclides, Montagner, Barbosa, do Valle and Nantes2016) found values of 0.640 kg LW/day for ADG and 3.8 AU/ha for SR when evaluating Piatã palisade grass during the rainy season (October to April). In the annual average (considering the dry season and rainy season), the authors found ADG and SR values of 0.428 kg LW/d and 2.43 AU/ha, respectively. The ADG and SR found in the present study can be considered enough for a period with low water availability (Fig. 1) that demonstrates the great potential that ICLS has to increase area use efficiency.
Furthermore, when ICLS is associated with nitrogen fertilization in the pasture phase, there is a significant response in the entire system. Higher soybean yields in the short term and this scenario reflects in improvements in soil biochemical and biological properties, as mentioned in Pires et al. (Reference Pires, Denardin, Silva, Freitas, Gonçalves, Camargo, Bremm, Carvalho and Souza2022), increase in GPA. This demonstrates that N fertilization in the pasture phase is essential to increase food production in the same area in ICLS.
Conclusions
The system fertilization strategy in Integrated Crop-Livestock Systems promotes greater accumulation and chemical composition of forage and, consequently, allows an increase in animal productivity. Alternatively, conventional fertilization with N application in the pasture phase in ICLS can be used to increase animal productivity. Pasture fertilization with P and K, without nitrogen fertilization in the pasture phase, increases weight gain per area due to higher stocking rate obtained.
Author contributions
CM Freitas: conceptualization, methodology, writing, investigation; JI Yasuoka: supervision, writing, data curation, formal analysis; GC Pires, JP Gama, LGS Oliveira and JEA Davi: writing, visualization, resources; LS Silva: project administration, supervision, data curation, writing-review-editing; IAG Silva: writing-review-editing, visualization; C Bremm: writing, investigation, methodology; PCF Carvalho: funding acquisition, project administration, supervision; A Moraes: funding acquisition, project administration, supervision; ED Souza: funding acquisition, project administration, supervision.
Funding statement
We thank the Fundação Agrisus, Yara Brasil, and REM-MT/FUNBIO for financial support. The funders had no role in data collection, preparation of the manuscript, or decision to submit for publication.
Competing interests
None.
Ethical standards
Not applicable.







