



Introduction
The Brazilian continental shelf presents a relative stability in physical and chemical parameters (Castro and Miranda, Reference Castro Filho and Miranda1998; Rossi-Wongtschowski and Madureira, Reference Rossi-Wongtschowski and Madureira2006; Mahiques et al., Reference Mahiques, Sousa, Furtado, Tessler, Toledo, Burone, Figueira, Klein, Martins and Alves2010). Despite the stability, this environment presents a high biodiversity (Rossi-Wongtschowski and Madureira, Reference Rossi-Wongtschowski and Madureira2006) due to high connectivity between the estuarine and shelf ecosystems for some fish species (Ibañez et al., Reference Ibañez, Hernández-Fraga and Alvarez-Hernández2017; Soeth et al., Reference Soeth, Spach, Daros, Adelir-Alves, Almeida and Correia2019; Kikuchi et al., Reference Kikuchi, Cardoso, Canel, Timi and Haimovici2021). Thus, this leads to high functional, physiological, and morphological plasticity for some species (Mai et al., Reference Mai, Condini, Albuquerque, Loebmann, Saint'Pierre, Miekeley and Vieira2014; Avigliano et al., Reference Avigliano, Leisen, Romero, Carvalho, Velasco, Vianna, Barra and Volpedo2017).
Otoliths are metabolically inert structures composed of calcium carbonate, mainly precipitated in the form of aragonite and countless chemical elements (Cadrin and Friedland, Reference Cadrin and Friedland1999; Campana, Reference Campana1999; Schulz-Mirbach et al., Reference Schulz-Mirbach, Ladich, Plath and Heb2019). The precipitation of calcium carbonate is regulated by hormones and influenced by environmental factors such as temperature and salinity (Popper and Fay, Reference Popper and Fay2011). The morphology of otoliths is complex, being influenced by growth parameters (Xiong et al., Reference Xiong, Yang, Zhang, Liu, Jiang and Chen2015), which could be related to morphological and morphometric variations such as distinct growth rates in the same population (Tuset et al., Reference Tuset, Jurado-Ruzafa, Otero-Ferrer and Santamaría2019), first maturation (Carvalho et al., Reference Carvalho, Volpedo and Fávaro2020), and physiological stress (Holmberg et al., Reference Holmberg, Wilcox-Freeburg, Rhyne, Tlusty, Stebbins, Nye, Honig, Johnston, San Antonio, Bourque and Hannigan2018). It is possible to describe patterns of habitat use by using ontogenetic variations in the otolith shape (Carvalho et al., Reference Carvalho, Vaz-dos-Santos, Spach and Volpedo2015; Bot et al., Reference Bot, Carvalho, Schwarz-Júnior and Spach2020). Conversely, widely distributed species may have different otolith morphotypes, as observed for Micropogonias furnieri (Santos et al., Reference Santos, Azevedo, Albuquerque and Araújo2017) and Trachurus picturatus (Vasconcelos et al., Reference Vasconcelos, Vieira, Sequeira, González, Kaufmann and Gordo2018).
The Sciaenidae family is composed by 270 species with a wide range of sizes (10–200 cm) (Chao, Reference Chao1978; Chao et al., Reference Chao, Frédou, Haimovici, Peres, Polidoro, Raseira, Subirá and Carpenter2015). Most of them live in shallow water (<50 m) (Odell et al., Reference Odell, Adams, Boutin, Collier, Deary, Havel, Johnson, Midway, Murray, Smith, Wilke and Yuen2017), with long or short life cycles of up to 31 years (Waggy et al., Reference Waggy, Brown-Peterson and Peterson2006; Cardoso and Haimovici, Reference Cardoso and Haimovici2011; Morat et al., Reference Morat, Marschal, Dominici and Harmelin-Vivien2017). Several species of Sciaenidae are commercially exploited and show complex migratory patterns (Albuquerque et al., Reference Albuquerque, Miekeley, Muelbert, Walther and and Jaureguizar2012; Childs et al., Reference Childs, Cowley, Næsje and Bennett2015). In the Southwestern Atlantic Ocean, the annual landings of sciaenids represent 22% of the total marine landings on the Brazilian coast (Chao et al., Reference Chao, Frédou, Haimovici, Peres, Polidoro, Raseira, Subirá and Carpenter2015). Larimus breviceps (Cuvier, 1830) is a demersal species widely distributed in the Central and Southwestern Atlantic (from Costa Rica to Santa Catarina in Brazil) (Vianna and Almeida, Reference Vianna and Almeida2005; Cattani et al., Reference Cattani, Santos, Spach, Budel and Gondim-Guanais2011). It has benthophagous habits and can attain up to 31 cm in total length (TL) (Santos et al., Reference Santos, Vasconcelos-Filho, Lira, Soares, Eduardo, Passarone, Le-Loc'h and Lucena-Frédou2021a; Froese and Pauly, Reference Froese and Pauly2023). The size at first maturity is known only for northeastern Brazil (11 cm TL, according to Santos et al., Reference Santos, Vasconcelos-Filho, Lira, Soares, Eduardo, Passarone, Le-Loc'h and Lucena-Frédou2021b). It has an important role in the food chain as prey of mammals and seabirds (Beneditto, Reference Beneditto2017; Miotto et al., Reference Miotto, Carvalho and Spach2017), and it is an abundant and frequent species of the bycatch of the coastal fisheries in Brazil, such as beach seine (Vianna and Almeida, Reference Vianna and Almeida2005; Passarone et al., Reference Passarone, Aparecido, Eduardo, Lira, Silva, Justino, Silva and Frédou2019) and shrimp trawling (Barreto et al., Reference Barreto, Freire, Reis, da Rosa, Carvalho Filho and Rotundo2018). This study aims at verifying the existence of any distinction in the shape of L. breviceps otoliths that may represent distinct morphotypes or stocks in three areas of the Brazilian coast. Thus, the results of this study can be used as a basis for future policies towards the management of different fish stocks.
Materials and methods
Study area
L. breviceps specimens were acquired in three localities along the Brazilian coast (Figure 1). The sampled specimens from northeastern Brazil were obtained from the commercial catches of shrimp trawlers based in Aracaju, state of Sergipe (10°55′41.07″S–37°02′34.08″W) (Figure 1A). Northeastern Brazil is characterized by bottom water temperatures ranging between 25.5 and 29.6°C (Cardoso and Haimovici, Reference Cardoso and Haimovici2011), being considered an oligotrophic environment (Heileman, Reference Heileman2009). This coast is subjected to a marked period of high precipitation in the austral winter, with an annual precipitation varying between 1000 and 1500 mm (Ekau and Knoppers, Reference Ekau and Knoppers1999).

Figure 1. Sampling sites of L. breviceps: (A) Aracaju – Sergipe, northeastern Brazil; (B) São Sebastião and Peruíbe – São Paulo, southeastern Brazil; and (C) Pontal do Paraná and Matinhos – Paraná, southern Brazil.
In the continental shelf of the southeastern region, the specimens were collected by gillnetters in the coast of the state of São Paulo (Peruíbe: 24°19′2″S–46°59′44″W; and São Sebastião: 23°45′40″S–45°24′44″W) (Figure 1B). In the coast of the state of Paraná, in southern Brazil, a subtropical environment, the specimens were collected by gillnetters in shallow coastal areas dominated by waves (Pontal do Paraná: 25°35′43.91″S–48°22′23.74″W; and Matinhos: 25°49′30.80″S–48°31′48.30″W) (Figure 1C). Southeastern–southern Brazil has lower temperature (sometimes below 21°C) and higher productivity, influenced by the constant nutrient input from estuaries (Rossi-Wongtschowski and Madureira, Reference Rossi-Wongtschowski and Madureira2006; Lessa et al., Reference Lessa, Santos, Souza Filho, Corrêa-Gomes, Lana and Bernardino2018) and by the intrusion of cold waters across the shelf (Central South Atlantic Water) with higher precipitation in the summer (Oliveira et al., Reference Oliveira, Aguiar, Cirano, Genz and Amorim2018). Its hydrography has wind and ocean currents as the main forces acting on the dynamics of the continental shelf, influencing the process of upwelling and intrusion of colder, nutrient-rich water masses that occur in the austral summer (Araújo and Gasalla, Reference Araújo and Gasalla2022). A water mass resulting from the mixture of coastal waters coming from the La Plata River, reinforced by the Patos Lagoon flow (Subtropical Shelf Water, SSW), flows over the platform extending to southeastern Brazil, being intensified by winds from the south quadrant in the winter. On the contrary, in the summer, the SSW converges with the coastal branch of the Brazilian Current, carrying tropical oligotrophic waters (Silveira et al., Reference Silveira, Schmidt, Campos, Godoi and Ikeda2000). This implies an alternation of these water masses on the continental shelf of southeastern and southern Brazil, which favours the larval dispersion of several fish resources, both northwards and southwards (Porcaro et al., Reference Porcaro, Zani-Teixeira, Katsuragawa, Namiki, Ohkawara and Favero2014).
Sample processing
After sampling, fish specimens were taxonomically identified, measured in TL (in cm, from the snout to the margin of the tail), and weighed in total weight (in g). Their sex was characterized macroscopically based on Vazzoler (Reference Vazzoler1996). Only adults of L. breviceps were used with a TL ranging between 13.8 and 20.0 cm in Sergipe (SE), 19.2 and 24.8 cm in São Paulo (SP), and 16.5 and 24.6 cm in Paraná (PR). All specimens used in the analyses presented length above the length at first maturity (Silva et al., Reference Silva, Viana, Fredou and Fredou2015; Santos et al., Reference Santos, Vasconcelos-Filho, Lira, Soares, Eduardo, Passarone, Le-Loc'h and Lucena-Frédou2021b). The sagitta otoliths were extracted from each fish, cleaned, packed dry, and numbered according to the geographic location.
Otolith contour analysis
The right otolith was photographed and the otolith length (OL, in mm) and otolith height (OH, in mm) were measured from these images (Figure 2A). The wavelet function was used to define the otolith contour (Parisi-Baradad et al., Reference Parisi-Baradad, Manjabacas, Lombarte, Olivella, Chic, Piera and García-Ladona2010; Sadighzadeh et al., Reference Sadighzadeh, Valinassa, Vosugi, Motallebi, Fatemi, Lombarte and Tuset2014) (Figure 2B). The wavelet is the result of the expansion of a signal in a family of functions representing expansions and translations of a mother function, i.e.: Ψs(x) = 1/sΨ(φ/s), where Ψ is a function with local support in a limited amplitude on the abscissa axis; φ lowers the step filter; and s is a scale parameter (Mallat, Reference Mallat1991). A total of 512 points, with equidistant coordinates from each otolith, were extracted with the rostrum as the contour origin (Parisi-Baradad et al., Reference Parisi-Baradad, Manjabacas, Lombarte, Olivella, Chic, Piera and García-Ladona2010). The fourth and fifth wavelets are more appropriate for identifying stocks or populations, as they describe the contour of the otoliths more sensitively (Sadighzadeh et al., Reference Sadighzadeh, Valinassa, Vosugi, Motallebi, Fatemi, Lombarte and Tuset2014). The image processing was performed using AFORO (http://aforo.cmima.csic.es/) (Parisi-Baradad et al., Reference Parisi-Baradad, Manjabacas, Lombarte, Olivella, Chic, Piera and García-Ladona2010).

Figure 2. (A) Otolith sagitta of L. breviceps. R, rostrum; A, anterior; D, dorsal; P, posterior; V, ventral region of the otolith; OL, otolith length; OH, otolith height. (B) Otolith contour using 512 equidistant points.
A principal component analysis (PCA), based on the variance–covariance matrix, was applied to reduce wavelet functions without losing information (Tuset et al., Reference Tuset, Imondi, Aguado, Otero-Ferrer, Santschi, Lombarte and Love2015). The principal components (PCs) explaining most data variability were selected by the broken-stick method (Gauldie and Crampton, Reference Gauldie and Crampton2002). Subsequently, the effect of the allometry of fish size was removed using the residuals of the linear regression between the significant PCs and the OL. From these, a new PCA was run (Stransky and MacLellan, Reference Stransky and Maclellan2005) to check for variations in the otolith contour for each study site: Sergipe, São Paulo, and Paraná. A linear discriminant analysis (LDA) was applied between sites to verify the correct percentage of otolith reclassification. A multivariate analysis of variance (MANOVA) was performed with the PCs without the effect of allometry, to check for differences in the shape of otoliths collected in those three states. All statistical analyses were performed using Past software version 4.03 (Hammer et al., Reference Hammer, Harper and Ryan2001).
Results
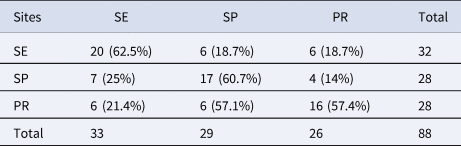
A total of 88 otoliths of L. breviceps from Sergipe, São Paulo, and Paraná were analysed (Table 1). The reconstruction of the otolith contour using wavelets 4 and 5 showed high variability in the contour of the specimens from the three states. This variability was observed in the dorsal, ventral, and posterior regions. Wavelet 4 showed greater variation between Sergipe and São Paulo–Paraná in the anterior region of the otoliths (Figure 3A). Wavelet 5 presented variation between Sergipe and São Paulo–Paraná in all regions of the otoliths (Figure 3B).
Table 1. Mean and standard deviation of fish TL, OL, and OH of L. breviceps by location and ‘n’ number of specimens analysed in Sergipe (SE), northeastern Brazil, in São Paulo (SP), southeastern Brazil, and in Paraná (PR), southern Brazil


Figure 3. Contour decomposition of the sagitta otolith of L. breviceps collected in Sergipe (SE), northeastern Brazil, in São Paulo (SP), southeastern Brazil, and in Paraná (PR), southern Brazil: (A) wavelet 4 and (B) wavelet 5.
The PCA showed high variability in the otolith shape (Figure 4). PC1 explained 97.6% and PC2 explained 1.7% of the otolith shape variability. Along PC1 more elongated otoliths are distributed with entire all margins, and on PC2 more rounded otoliths are distributed with entire all margins, whose anterior region presents a prominence and slight excisura. Otoliths of L. breviceps from SP and PR were more distributed along PC1 with more elongated otolith shape and those from SE were differentiated from them along PC2 with more rounded otoliths (Figure 4).

Figure 4. PCA scatterplot for the sagitta otolith contour of L. breviceps from Sergipe (SE; yellow dots), northeastern Brazil, São Paulo (SP; green dots), southeastern Brazil, and Paraná (PR; blue dots), southern Brazil.
The LDA showed correct reclassification of 60.23% of all otoliths among sites. SE otoliths showed the highest reclassification percentage (62.50%), followed by PR (60.71%) and SP (57.14%) (Table 2). The MANOVA evidenced a significant difference in otolith shape among sites (F = 2.3; P < 0.05). The otolith shape from specimens originating from SE was significantly different from those originating from SP (P < 0.05).
Table 2. Reclassification of sagittae otolith of L. breviceps between those collected in the northeastern (Sergipe, SE), southeastern (São Paulo, SP), and southern Brazil (Paraná, PR) by the LDA
Discussion
The sagitta otoliths of L. breviceps presented a bullet shape, characteristic of many sciaenids (Volpedo and Echevarria, Reference Volpedo and Echeverría1999; Waessle et al., Reference Waessle, Lasta and Favero2003; Tuset et al., Reference Tuset, Lombarte and Assis2008; Siliprandi et al., Reference Siliprandi, Rossi-Wongtschowski, Brenha, Gonsales, Santificetur and Vaz-dos-Santos2014). Two morphotypes were identified, the most elongated in northeastern Brazil (SE) and one with prominence and excisura in southeastern–southern Brazil (SP–PR). The presence of two morphotypes may indicate environmental influences and connectivity (or not) between populations.
Northeastern Brazil is characterized by a bottom water temperature varying between 25.5 and 29.6°C (Cardoso and Haimovici, Reference Cardoso and Haimovici2011), and it is considered an oligotrophic environment (Heileman, Reference Heileman2009). Conversely, southeastern–southern Brazil presents lower temperature and higher productivity, influenced by constant input of nutrients from estuaries (Rossi-Wongtschowski and Madureira, Reference Rossi-Wongtschowski and Madureira2006; Lessa et al., Reference Lessa, Santos, Souza Filho, Corrêa-Gomes, Lana and Bernardino2018). These environmental differences may have influenced the presence of two morphotypes in the otoliths of L. breviceps. Other sciaenids also showed distinct morphotypes influenced by habitat and environmental parameters. Paralonchurus brasiliensis (Steindachner, 1875) e.g. presented elongated morphotype otoliths as a result of a river nutrient input, differing from the rounded morphotype found in populations living far from the mouth of the Paraíba do Sul River (Oliveira et al., Reference Oliveira, Di Beneditto and Monteiro2009). Argyrosomus japonicus (Temminck & Schlegel, 1843) also presented two morphotypes, associated with the presence of a river input and, hence, higher productivity influencing the growth rate (Ferguson et al., Reference Ferguson, Ward and Gillanders2011). M. furnieri (Desmarest, 1823) presented three otolith morphotypes along southeastern–southern Brazil, related to oceanographic processes occurring far from the coast (Santos et al., Reference Santos, Azevedo, Albuquerque and Araújo2017). Thus, more productive areas provide greater food availability influencing the growth rate of otoliths and their morphology (Verocai et al., Reference Verocai, Lombarte and Norbis2023).
The continental shelf of southeastern–southern Brazil represents a transitional region between tropical and subtropical climates and has 13 large estuaries increasing the input of nutrients to the continental shelf (Spalding et al., Reference Spalding, Fox, Allen, Davidson, Ferdaña, Finlayson, Halpern, Jorge, Lombana, Lourie, Martin, McManus, Molnar, Recchia and Robertson2007; Lessa et al., Reference Lessa, Santos, Souza Filho, Corrêa-Gomes, Lana and Bernardino2018). This environmental similarity between southeastern and southern Brazil (SP and PR) is probably the reason behind the similar shapes of L. breviceps otoliths sampled at those areas. Along with the similarity of environmental parameters, the otolith shape may indicate connectivity between populations of L. breviceps from São Paulo and Paraná. L. breviceps inhabits the shallow inner shelf (<50 m deep) in all ontogenetic phases (Bessa et al., Reference Bessa, Santos, Pombo, Denadai, Fonseca and Turra2014; Porcaro et al., Reference Porcaro, Zani-Teixeira, Katsuragawa, Namiki, Ohkawara and Favero2014), allowing gene flow between populations of L. breviceps, resulting in a similar otolith shape between these two populations. However, the morphotype of otoliths is influenced by several parameters, such as temperature and salinity (Popper and Fay, Reference Popper and Fay2011). In addition to environmental parameters, genetics and reproductive behaviours can influence shape otoliths. Recent studies demonstrate the importance of genetics and otolith shape in populational studies of marine fishes. Results of Ibanez et al. (Reference Ibañez, Rangely, Avila-Herrera, Silva, Almanzar, Neves, Avigliano, Callicó, Volpedo and Fabré2022), e.g. demonstrated that genetics studies for Mugil curema indicated variations on a macro-scale and otoliths on smaller scales. A similar otolith shape between populations having gene flow has been observed in other species, such as the Batrachoididae Opsanus beta (Goode & Bean, 1880) (Carvalho et al., Reference Carvalho, Martinez-Pérez, Aguilar-Perera, Quiñones, Tomás, Vitule and Volpedo2022), and the Sciaenidae Umbrina canosai Berg, 1895 (Kikuchi et al., Reference Kikuchi, Cardoso, Canel, Timi and Haimovici2021).
The analysis of the contour of otolith shape is an important tool which suggested the existence of connected and separated populations of L. breviceps along the Brazilian coast. We further recommend studies involving the otolith chemistry and genetics to assess possible migration between the populations of the northeastern and southeastern–southern Brazil.
Data availability
The data that support the findings of this study are available from the corresponding author upon request.
Acknowledgements
B. M. C. is thankful to the National Council for Scientific and Technological Development (CNPq no. 168196/2022-0). A. R. G. T. is thankful to Sao Paulo Research Foundation (FAPESP no. 2018/04099-5). K. M. F. F. and B. A. S. thank Diógenis Lopes for helping to obtain the samples at the landing port of Aracaju – Sergipe, and Jessica Diniz, Tatiana da Silva, and Izabela Canuto for helping with sample processing at the laboratory.
Author contributions
Barbara Maichak de Carvalho – study design, sampling, analysis, writing, and revision. Yasmin Barbieri – sampling and writing. Beatriz Andrade Syrio – sampling and writing. Kátia Meirelles Felizola Freire – sampling, writing, and revision. Acácio Ribeiro Gomes Tomás – sampling, writing, and revision.
Financial support
This study was supported by National Council for Scientific and Technological Development (CNPq no. 168196/2022-0).
Competing interests
None.






