


Implications
The use of chicory within ruminant systems is increasing due to the benefits shown for this forage in sheep production systems. However, there has been little research on the effects of using chicory in beef systems. Here, the effects of chicory inclusion in the diet of grazing steers on meat quality and sensory properties were investigated. Findings showed there were no substantive effects of including chicory in the swards of grazing beef cattle on meat stability, fatty acid composition or sensory properties of the M. Longissimus muscle when compared with beef steers grazing ryegrass-only swards.
Introduction
Chicory (Cichorium intybus) has been regarded for many years worldwide as a valuable constituent of pastures for grazing livestock. As the 1980s onwards, with the development of commercial forage cultivars (Rumball et al., Reference Rumball, Skipp, Keogh and Claydon2003), the use of chicory has been steadily increasing albeit predominately in sheep and deer production systems. Research findings have confirmed the attributes for this forage include its ability to increase the productivity of finishing lambs and reduce internal helminth parasites (Marley et al., Reference Marley, Barrett, Lampkin, Cook and Keatinge2003). The livestock production attributes from chicory are mostly due to its high CP and energy values as well as it being highly productive agronomically resulting in an improvement in the seasonal availability of high quality forage (Li and Kemp, Reference Li and Kemp2005).
Chicory leaves contains a range of secondary plant compounds, including sesquiterpene lactones (bitter compounds), tannins and other phenolic compounds (Kisiel and Zielińska, Reference Kisiel and Zielińska2001). These dietary plant secondary metabolites have been shown in numerous studies to affect meat quality (Priolo et al., Reference Priolo, Micol and Agabriel2001). Most notably, degradation products of sesquiterpene lactones found in chicory, namely dihydrolactucin, tetrahydrolactucin and hydroxypenyacetic acid, were identified as the taint compounds in the milk of dairy cows on high chicory diets (Visser, Reference Visser1992), leading to speculation that the same compounds could result in meat with a bitter taste compared with meat from grass-based systems. In a recent study, consumers preferred steaks from steers finished on pearl millet (Pennisetum glacum), cowpea (Vigna unguiculata) or lucerne (Medicago sativa) when compared with steaks from steers grazing chicory or bermudagrass (Cynodon dactylon) (Schmidt et al., Reference Schmidt, Miller, Andrae, Ellis and Duckett2013).
Consumers are increasingly aware of the importance of diet on their health and this has resulted in consumer interest in the nutritional value of foods (Scollan et al., Reference Scollan, Dannenberger, Nuernberg, Richardson, MacKintosh, Hocquette and Maloney2006). In relation to the healthiness of meat, the industry is largely concerned with the total amount of fat and its fatty acid composition, aiming to produce meat with a higher ratio of polyunsaturated fatty acids (PUFA) to saturated fatty acids and the optimal ratio of n-6 and n-3 PUFA (Wood et al., Reference Wood, Richardson, Nute, Fisher, Campo, Kasapidou, Sheard and Enser2004). Fatty acid profiles have been shown to differ among forages, with chicory reported as having a different profile and a higher total concentration of PUFA (C18 : 3n-3) than other forages including ryegrass (Clapham et al., Reference Clapham, Foster, Neel and Fedders2005). In the experiment by Schmidt et al. (Reference Schmidt, Miller, Andrae, Ellis and Duckett2013), chicory increased the ratio of n-6/n-3 PUFA in the meat of grazing steers when compared with steers grazing bermudagrass, cowpea or lucerne, suggesting that including chicory in the diet of grazing ruminants would alter the total fatty acid concentration and composition of their meat.
Despite the production benefits highlighted for chicory within ruminant diets and research suggesting that the fatty acids and other secondary compounds present in this forage may alter meat properties when consumed by farmed animals, there have been no studies into the effects of this forage when compared with perennial ryegrass swards on beef quality within a temperate pasture-based beef finishing system. The aim of the experiment described here was to determine the effects of chicory/perennial ryegrass swards compared with perennial ryegrass swards on the meat stability, fatty acid composition and sensory properties of the M. Longissimus muscle of grazing beef steers grazing ryegrass.
Material and methods
Experimental site, animals and forage treatments
Full details of the grazing experiment, including plot establishment, grazing management and performance data are presented in Marley et al. (Reference Marley, Fychan, Davies, Scollan, Richardson, Theobald, Genever, Forbes and Sanderson2014). In brief, triplicate 2 ha field plots were established with either a chicory (cv. Puna II)/perennial ryegrass (cv. Premium) mix or a perennial ryegrass control (cv. Premium) at Penglais farm, Aberystwyth University (52°25'46'N 4°4'13'W). The experiment was conducted over 2 consecutive grazing years (2010–11), with individual animals remaining on the same replicate forage plot in both years. A core group of 36 Belgian Blue – dairy cross steers (mean±SEM live weight: 184±4.7 kg aged ~7 months on Day 0) were retained for the whole 2-year production and finishing period, resulting in six animals per replicate plot. The measurement period ran from 25 May until 28 September and 12 April until 11 October, in the 1st and 2nd grazing year. In the winter period between grazing seasons, animals were housed as one group and offered a standard diet of ryegrass silage plus straw. In the 2nd grazing year, steers were selected for slaughter as they were deemed to have reached a fat class of 3, according the EU regulations EUROP classification (where 1 is very lean and 5 is very fat) (MLCSL, 2016). At this stage, steers from each treatment group were transported on the day before slaughter from the Institute of Biological, Environmental and Rural Sciences at Aberystwyth University to the Food Science and Food Safety Group at the University of Bristol, Langford, UK. All animal procedures followed strict guidelines as set forward in the Animals (Scientific Procedures) Act (1986) and approved by the Home Office (HO), UK and were performed under a HO licence. Animals were certified as not suffering any adverse effects as a result of any procedures at the end of the experimental period by the named HO veterinary officer.
Sample collection
Animals were housed overnight before slaughter. Animals were stunned with a captive bolt gun and killed by severing the carotid arteries. Carcasses were subjected to electrical stimulation using a low voltage electrical stimulation unit (MIRINZ, Hamilton, New Zealand) delivering 90 v (125 to 500 mA, pulse width 5 to 10 ms, pulse period 57 to 80 ms (14.3 Hz)) for 60 s, 30 s after bleeding began. After dressing, half carcass sides were hung in a 2°C chiller for 48 h and then then quartered between the junction of the 10th and 11th rib. A 250 mm-long section of the hindloin joint containing the M. Longissimus lumborum muscle was removed from the left side of the carcass, posterior to the 10th rib. A 20 mm-thick steak was cut and the muscle dissected free of subcutaneous adipose tissue. The muscle sample was vacuum packed and frozen at −20°C for subsequent analysis of vitamin E, a further steak being retained for fatty acid analysis. The remaining section of the loin was vacuum packed and conditioned at 1°C to 14 days from slaughter. After this, three 20 mm-thick steaks were cut and packed individually in modified atmosphere (MA) packs (O2 : CO2; 75 : 25) and subjected to simulated retail display (4°C, 700 lux for 16 h out of 24 h). The remaining section of the conditioned loin was frozen and stored at −20°C before sensory analysis.
Meat and fat measurements
Muscle pH was checked at 2 and 48 h post-slaughter in the M. Longissimus between the 10th and 11th rib, using a Testo 230 pH direct probe (Testo, Wilmington, NC, USA), calibrated with pH 4.0 and 7.0 buffers. Fat colour (L*a*b*) was determined on the cold carcass at 48 h post-slaughter on the subcutaneous fat over the 10th rib, 10 cm from the mid-line avoiding any blood streaked areas using a Minolta CR400 Chromometer (Minolta UK Ltd, Milton Keynes, UK) standardised as specified by Cassens et al. (Reference Cassens, Demeyer, Eikelenboom, Honikel, Johansson, Nielson, Rennerre, Richardson and Sakata1995) on a white tile. The L*a*b* system of the Commission International de L’Eclairage (Commission Internationale de l’Eclairage, 1978) was used and values for hue (tan−1 b*/a*) and saturation (square root of (a*2+b*2) calculated. Meat colour was measured daily on two MA packed steaks on three areas of the surface of the steaks through the film lid and averaged, until the colour changed from red to brown. Two further MA packed steak was taken at day 10, trimmed of excess visible fat around the edges, homogenised and subsampled, for analysis of lipid oxidation as thiobarbituric acid reacting substances (TBARS) by the method of Tarladgis et al. (Reference Tarladgis, Watts, Younathan and Dugan1960) modified by the use of a Buchi 321 distillation unit (BUCHI Corporation, New Castle, DE, USA). Vitamin E was analysed by the method of Liu et al. (Reference Liu, Scheller and Schaefer1996), using 5,7-dimethyl-tocol as internal standard.
Fatty acid composition and profile
Fatty acids analysis was carried out by direct saponification as described in detail by Teye et al. (Reference Teye, Sheard, Whittington, Nute, Stewart and Wood2006). Samples were hydrolysed with 2 M KOH in water:methanol (1 : 1) and the fatty acids extracted into petroleum spirit, methylated using 14% BF3 in methanol and analysed by GLC. Samples were injected in the split mode, 70 : 1, onto a CP Sil 88, 50 m×0.25 mm fatty acid methyl esters (FAME) column (Chrompack UK Ltd, London, UK) with helium as the carrier gas. The output from the flame ionization detector was quantified using a computing integrator (Spectra Physics 4270, Santa Clara, CA, USA) and linearity of the system was tested using saturated (FAME4) and monounsaturated (FAME5) methyl ester quantitative standards (Thames Restek UK Ltd, Windsor, Universal Biologicals (Cambridge) Ltd., Cambridge, UK), PUFA (n-3; Matreya, Universal Biologicals (Cambridge) Ltd., Cambridge, UK), short to medium chain and branched chain fatty acids (bacterial acid methyl ester mix; Supelco, Sigma-Aldrich Company Ltd., Gillingham, UK) and a mix of C20 and C22 n-3 and n-6 fatty acids made in the laboratory from a mix of methyl esters (Sigma, Sigma-Aldrich Company Ltd., Gillingham, UK). Total intramuscular fat concentration was calculated gravimetrically as total weight of fatty acid extracted.
Sensory (eating) quality
Sensory analysis was carried out by a 10-person trained taste panel (BSI, 1993). The samples were defrosted overnight at 4°C and then cut into steaks 20 mm thick. Steaks were grilled to an internal temperature of 74°C (measured by a thermocouple probe), turning over every 2 min, after which all fat and connective tissue was trimmed and the muscle cut into 2 cm cubes. The blocks were wrapped in pre-labelled foils and placed in a heated incubator. The samples were then given to the assessors in a random order assigned by computer, according to MacFie et al. (Reference MacFie, Bratchell, Greenhoff and Vallis1989). Panellists assessed samples from carcasses of two different chicory-grazed and two grass-grazed animals per session, four sessions in a morning using an eight-point category scale for tenderness, juiciness, beef flavour and abnormal flavour and a 0 to 100 mm unstructured line scales where 0=nil and 100=extreme intensity for flavour descriptors chosen by the panel in previous training sessions.
Statistical analysis
All data were examined by ANOVA using GenStat® (Payne et al., Reference Payne, Murray, Harding, Baird and Soutar2014) and treating plots as experimental units and pasture type as a fixed effect. Colour saturation time courses were analysed by repeated measures ANOVA. Fatty acid composition as a percentage of the total fatty acid data were analysed by principal component analysis using Genstat®. Taste panel intensity ratings (either on an eight-point category scale or 0 to 100 scale, values were standardised for panellists with the Fizz software and compared by ANOVA treating plots as experimental units with pasture type as a fixed effect.
Results
Meat and fat measurements
Muscle pH measured in the Longissimus muscle at 2 and 48 h after slaughter, fat colour, muscle vitamin E concentration and muscle fat oxidation stability (shown as TBARS) are presented in Table 1. These data showed there were no effects (P>0.05) on meat quality of including chicory in the diet of grazing beef steers in the present study. As the display period in the modified atmosphere packs progressed, numerical difference in colour saturation decreased between the dietary treatment groups (Figure 1). However, in spite of numerically lower saturation values for samples from animals fed chicory at 24 h after packing in MA, and this difference persisting for 5 days post-packaging, there was no statistical evidence that diet influenced saturation or the rate of decline with time (P>0.05).
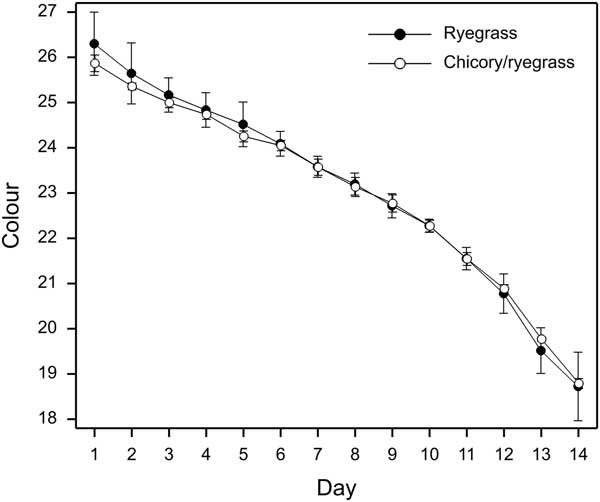
Figure 1 Effect of dietary treatment and time in simulated retail display in modified atmosphere packs on colour saturation (±SEM) of M. Longissimus steaks from steers grazing either chicory/ryegrass or ryegrass-only swards in a 2-year pasture-based finishing system.
Table 1 Effect of either chicory/ryegrass or ryegrass dietary treatments on the pH, temperature and stability of the M. Longissimus and carcass fat colour from beef steers grazing in a 2-year pasture-based finishing system

PRG=perennial ryegrass; CH/PRG=chicory/ perennial ryegrass; SED=standard error of the difference; Ns=not significant (P>0.05); TBARS=thiobarbituric acid reacting substances.
1 SED shown applies to means transformed to log10 scale.
Fatty acid composition and profile
Data showed there were no differences in the levels of the total amount or relative proportion of selected individual fatty acids in the M. Longissimus steaks taken from beef steers grazing chicory/ryegrass or ryegrass-only swards within a 2-year pasture-based finishing system (P>0.05) (Table 2). Figure 2 shows selected fatty acid compositions as a percentage of the total fatty acids did not differ between treatments (P>0.05).
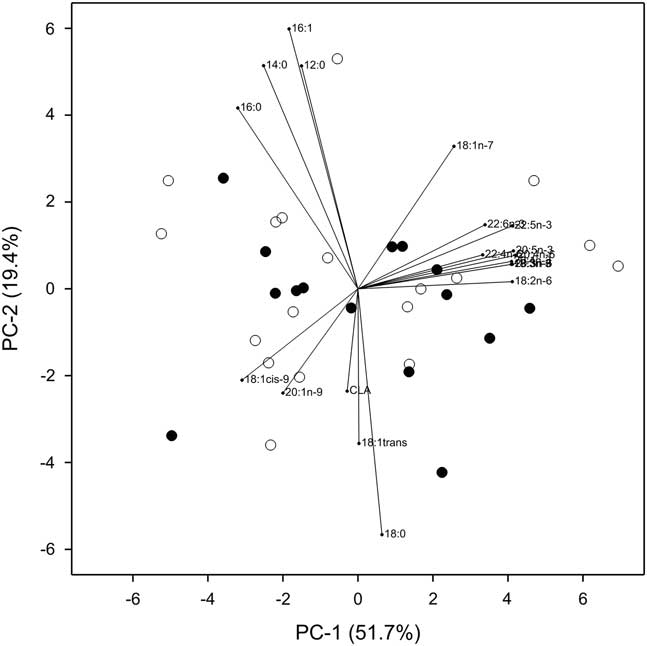
Figure 2 Principal component bi-plot showing the fatty acid compositions as a percentage of the total fatty acids of M. Longissimus steaks from steers grazing either chicory/ryegrass or ryegrass-only swards in a 2-year pasture-based finishing system.
Table 2 Effect of either chicory/ryegrass or ryegrass dietary treatments on the selected fatty acid concentration (mg/100 g tissue), total fatty acid concentration (mg/100 g tissue) and fatty acid ratios of the M. Longissimus from beef steers grazing in a 2-year pasture-based finishing system

PRG=perennial ryegrass; CH/PRG=chicory/ perennial ryegrass; SED=standard error of the difference; Ns=not significant (P>0.05); SFA=saturated fatty acids; MUFA=monounsaturated fatty acids; PUFA=polyunsaturated fatty acids; CLA=conjugated linoleic acid.
1 When using a 50-m CP-SIL 88 column, 18 : 1 trans-6 to trans-11 are detected as a single peak, and 18 : 1 trans-12, -13, -14, -15 and -16 and 18 : 1 cis-6 co-elute with 18 : 1 cis-9 (Molkentin and Precht, Reference Molkentin and Precht1995). Therefore, measurements of 18 : 1 trans-6 to trans-11 were used to estimate total trans 18 : 1 concentrations.
2 Sum of trans-12 to trans-16 18 : 1 was used to correct cis-9 18 : 1 concentrations.
3 18 : 2cis-9, trans-11 (may also contain some trans-8, cis-10 CLA and trans-7, cis-9 CLA as minor isomers).
4 SFA: sum of 12 : 0, 14 : 0, 16 : 0, 18 : 0.
5 MUFA: sum of 16 : 1, 18 : 1, 18 : 1n-9, 18 : 1n-7, 20 : 1n-9.
6 PUFA: sum of 18 : 2n-6, 18 : 3n-3, 20 : 3n-6, 20 : 4n-6, 20 : 4n-3, 20 : 5n-3, 22 : 4n-6, 22 : 5n-3, 22 : 6n-3.
7 n-3, sum of 20 : 4n-3, 20 : 5n-3, 22 : 5n-3, 22 : 6n-3.
8 n-6, sum of 18 : 2n-6, 20 : 3n-6, 20 : 4n-6, 22 : 4n-6.
Sensory (eating) quality
Taste panel assessments found the sensory qualities of beef steaks from beef steers grazing chicory/ryegrass or ryegrass-only swards to be comparable with no difference in eating qualities, flavours or overall liking (P>0.05) (Table 3).
Table 3 Effect of either chicory/ryegrass or ryegrass dietary treatments on the taste panel intensity ratings (either on an eight-point category scale or 1 to 100 scale, as stated) for beef steaks from steers grazing in a 2-year pasture-based finishing system.

PRG=perennial ryegrass; CH/PRG=chicory/ perennial ryegrass; SED=standard error of the difference; Ns=not significant (P>0.05).
1 Eight-point category scale used where 1 to 4=extremely, very, moderately, slightly lacking in the trait; 5 to 8=slightly, moderately, very and extremely present in the trait.
2 Hedonic assessment.
Discussion
The main findings of current study was that there were no substantive effects of including chicory in the swards of grazing beef cattle on meat stability, fatty acid composition or sensory properties when compared with beef steers grazing ryegrass-only swards. Many factors influence ruminant meat quality and these can be divided into two main categories: the animal factors (e.g. breed, age, gender) and environmental factors (e.g. diet, weather, slaughtering procedures) (Priolo et al., Reference Priolo, Micol and Agabriel2001). In many studies, the effect of diet on meat quality can be confounded by the effects of the diet on growth rates, making it difficult to determine if the meat characteristics are due to the effects of the diet or its effects on body composition per se (Muir et al., Reference Muir, Deaker and Bown1998). In the study reported here, previously published data confirmed that there were no differences in the growth rates, final live weight or carcass characteristics of the beef steers grazing either dietary treatment (Marley et al., Reference Marley, Fychan, Davies, Scollan, Richardson, Theobald, Genever, Forbes and Sanderson2014), allowing for a clear interpretation of the data without these factors confounding the investigation.
Meat and fat measurements
The colour saturation during retail simulation within the current study were not found to differ with both treatment being within the typical range reported for grazing systems (Priolo et al., Reference Priolo, Micol and Agabriel2001) and remaining above 18 until day 14. A chroma value of 18 is considered the threshold value, which corresponds to the formation of 20% metmyoglobin (McDougall, Reference McDougall1982), the borderline value for consumer rejection during the purchase of fresh beef (Hood and Riordan, Reference Hood and Riordan1973). Beef colour stability is known to differ among muscle type, with the M. Longissimus muscle being the most stable (O’Keeffe and Hood, Reference O’Keeffe and Hood1982) and, therefore the findings reported here are only in reference to this muscle and may have differed if other muscle types were studied. In the current study, the muscle pH values, as shown by muscle pH values at 48 h, show that the meat pH from both treatments in the current study was within the normal range (pH 5.5 to 5.8) for beef (Priolo et al., Reference Priolo, Micol and Agabriel2001), including studies on the M. Longissimus muscle from pasture-based systems (Warren et al., Reference Warren, Scollan, Enser, Hughes, Richardson and Wood2008a).
The concentration of vitamin E in meat affects muscle pigment in the longissimus muscle (myoglobin to brown metmyoglobin) and lipid oxidation (Realini et al., Reference Realini, Duckett, Brito, Dalla Rizza and De Mattos2004). Studies have shown that chicory contains significantly higher concentrations of δ-tocopherol (16.6 mg/kg dry matter (DM)) when compared with other forages (Larsen et al., Reference Larsen, Fretté, Kristensen, Eriksen, Søegaard and Nielsen2012) but there were no differences in concentrations of the more bioactive forms of α- or β-tocopherol. Further research is needed to determine the effects of different tocopherol isomers in beef diets on meat quality. In the current study, the vitamin E levels in the muscle of beef steers are similar when compared with those reported (3.91 μg/g as α-tocopherol) for the longissimus muscle of pasture fed beef by Realini et al. (Reference Realini, Duckett, Brito, Dalla Rizza and De Mattos2004) and higher than those reported for the longissimus muscle silage-fed beef (Warren et al., Reference Warren, Scollan, Enser, Hughes, Richardson and Wood2008a). Therefore, a lack of difference in the meat vitamin E concentrations between the two treatments may also have contributed to the finding that there was no differences in the rate of change of colour chroma during the simulated retail display period. The TBARS values found in the current study are within an acceptable range expected for the longissimus muscle of cattle grazing pastures, also likely due to elevated vitamin E concentrations (Warren et al., Reference Warren, Scollan, Enser, Hughes, Richardson and Wood2008a).
The inclusion or deficiency of certain minerals, which play a role in metalloproteins which act as antioxidants, such as copper (Huang et al., Reference Huang, Wang, Lin and Guo2014) and cobalt (Kennedy et al., Reference Kennedy, Kennedy, Blanchflower, Scott, Weir, Molloy and Young1994) and selenium have been shown to alter meat quality either by reducing the rate of myoglobin or lipid oxidation of the longissimus muscle in beef (Realini et al., Reference Realini, Duckett, Brito, Dalla Rizza and De Mattos2004), or lipid metabolism in tissues in sheep (Kennedy et al., Reference Kennedy, Kennedy, Blanchflower, Scott, Weir, Molloy and Young1994). Research has reported there are higher concentrations of copper and cobalt in chicory compared with ryegrass (Marley et al., Reference Marley, Fychan, Scott, Davies and Sanderson2013). However, data on the forage chemical composition (presented in Marley et al., Reference Marley, Fychan, Davies, Scollan, Richardson, Theobald, Genever, Forbes and Sanderson2014) showed no differences in ash concentration between the two forage treatments, which suggest the mineral composition may not have differed in the current study. However, the elevated vitamin E concentrations found in the meat in this study could have been responsible for the extended colour shelf life. Further studies are now needed to understand if there are any effects of the inclusion of chicory on the health value of the beef produced for human nutrition, such as higher levels of essential micro-nutrients or other vitamins.
Fatty acid composition and profile
It has been well defined in numerous studies that dietary fat concentration and fatty acid composition can alter meat quality (Wood et al., Reference Wood, Richardson, Nute, Fisher, Campo, Kasapidou, Sheard and Enser2004; Scollan et al., Reference Scollan, Hocquette, Nuernberg, Dannenberger, Richardson and Moloney2014). Increasing the PUFA concentration in the diet of cattle typically increases the concentration of these fatty acids in beef (Scollan et al., Reference Scollan, Choi, Kurt, Fisher, Enser and Wood2001). In general, differences in the fatty acid ratios in meat are most apparent in studies comparing contrasting forage-based diets against concentrate-based diets (Warren et al., Reference Warren, Scollan, Nute, Hughes, Wood and Richardson2008b) and could be one explanation as to why there was no differences in the current study, particularly as the n-6:n-3 ratio was as low as 1.05 for the control, with below 3 being typical for beef (Scollan et al., Reference Scollan, Dannenberger, Nuernberg, Richardson, MacKintosh, Hocquette and Maloney2006). That stated, fatty acid profiles have been shown to differ among forages, with chicory reported as having higher concentrations of C18 : 3 when compared with grasses (including ryegrass), plantain, turnips, rape, triticale and borage (Clapham et al., Reference Clapham, Foster, Neel and Fedders2005). In contrast to the current study, other research found that grazing beef steers on chicory increased the ratio of n-6 to n-3 fatty acids in the meat when compared with steers grazing bermudagrass, cowpea or lucerne but not those grazing pearl millet (Schmidt et al., Reference Schmidt, Miller, Andrae, Ellis and Duckett2013). Further research into the fatty acids profiles in chicory when grown and managed under different environmental conditions are now required to determine if feeding chicory to cattle could be used as an effective approach to improving beef fatty acid composition, with potentially beneficial consequences for human nutrition (Scollan et al., Reference Scollan, Dannenberger, Nuernberg, Richardson, MacKintosh, Hocquette and Maloney2006).
Sensory (eating) quality
Further physiological mechanisms by which chicory could exhort an effect on the quality of meat from ruminant livestock may be due to its phytochemical composition. Chicory leaves contains a range of secondary plant compounds, including sesquiterpene lactones (bitter compounds), tannins and other phenolic compounds (Kisiel and Zielińska, Reference Kisiel and Zielińska2001). These secondary metabolites have been shown scientifically to have effects on meat quality. Feeding diets with condensed tannins increases ultimate-pH in meat (Priolo et al., Reference Priolo, Waghorn, Lanza, Biondi and Pennisi2000). More notably, sesquiterpene lactones, found in chicory leaves (Kisiel and Zielińska, Reference Kisiel and Zielińska2001), may reduce skatole (3-methyl indole), one of the compounds widely considered to cause boar taint in pork meat. In other research, degradation products of these sesquiterpene lactones, namely dihydrolactucin, tetrahydrolactucin and hydroxypenyacetic acid, were identified as the taint compounds in the milk of dairy cows on high chicory diets (Visser, Reference Visser1992). These accounts led to anecdotal speculation that the same compounds could result in meat with a bitter taste. In the findings of the current study, there was no effect of including chicory in the diet of beef steers on eating quality. However, determining the concentrations of these secondary metabolites over the 2-year period of the current study, to confirm how these compare with other studies, was outside the constraints of the research so further research is needed to increase our understanding of the role of the secondary metabolites in chicory in meat quality.
Our findings are in contrast to the research comparing beef from steers grazing chicory to bermudagrass, cowpea, lucerne or pearl millet, where it was found that consumers preferred steaks from steers finished on all other treatments compared with chicory or bermudagrass (Schmidt et al., Reference Schmidt, Miller, Andrae, Ellis and Duckett2013). Although it is difficult to draw comparisons between cattle and sheep given species differences in grazing efficiencies and intakes of chicory (McCoy et al., Reference McCoy, Collins and Dougherty1997), research looking at pure chicory swards compared with ryegrass in finishing lamb systems found, in agreement with our study, that including chicory in the diet of grazing lambs did not have any substantive effects on meat eating quality (Houdijk et al., Reference Houdijk, Baker, Navajas, Richardson and Ross2011).
In practical farm situations, chicory is typically used in one of two ways: either as a medium-term ley in a mix with grass and clovers or as a pure sward for a short-term lamb or deer-finishing crop, with most of the research showing higher productivity responses for sheep and deer being conducted using the specialist pure short-term forage chicory crops. However, due to the long-term nature of forage-based beef finishing systems, in the current study the chicory was sown within a mix with perennial ryegrass (sward contained 24% and 14% chicory on a DM basis in Years 1 and 2, respectively; see Marley et al., Reference Marley, Fychan, Davies, Scollan, Richardson, Theobald, Genever, Forbes and Sanderson2014 for all forage data) to make the current experiment applicable to a practical farming system. Using chicory within a ryegrass mix was also more likely to reduce the risk of any potential effects of a wet grazing season on either DM intakes or sward damage whilst a 2-year experiment was completed. However, it is possible that this reduced the potential to detect any effects of chicory on meat quality that could result from offering a pure sward of chicory as the forage diet to grazing beef steers, either grazed each season or in the second season only, and further studies involving short-term grazing studies on pure swards of chicory are needed to investigate this.
Previous research into the effects of different forage species on meat quality, coupled with research showing that dietary fatty acid, antioxidant and secondary plant compounds can alter meat quality, provided some evidence that the inclusion of chicory in pasture swards might alter the meat colour stability, fatty acid and sensory properties of beef when used within pasture-based beef finishing systems. However, the overall findings from this current study showed that chicory can be offered to beef steers when used within ryegrass/chicory swards without any substantive effects on meat stability, fatty acid composition or sensory properties of the M. Longissimus muscle. Further studies into the effects of short-term pure chicory swards grazed either each season or in the second season only are now needed to further investigate the effects of using chicory in pasture-based beef finishing systems.
Acknowledgements
The authors would like to gratefully acknowledge the advice received on this work from Phil Evans (retired). The authors would like to gratefully acknowledge M. Leyland, N. Gordon, M. Scott, O. Martin, A. Baker, K.G. Hallett and H. Fleming for their help in sample collection and measurements. This work was sponsored by EBLEX (now AHDB Beef and Lamb), UK. Seed for the swards was donated by Germinal, UK. Institute of Biological, Environmental and Rural Sciences receives strategic funding from the Biotechnology and Biological Sciences Research Council.






 Open access
Open access