


Weeds can account for up to 31% of potential yield loss in agricultural crops and remain one of the most significant biotic factors reducing yield in both conventional and organic farming systems (Bond and Grundy Reference Bond and Grundy2001; Oerke Reference Oerke2006). However, conventional and organic systems differ significantly in their approaches to weed management. Conventional arable farming systems rely heavily upon herbicides, a strategy that has been incredibly effective at controlling weeds. However, such reliance on herbicides has resulted in both inappropriate use and overuse, the result of which can be the evolution of herbicide-resistant weed populations that become increasingly difficult to control (Heap Reference Heap2014). Furthermore, there has been concern for many years about the environmental and human health risks posed by production and application of herbicides (Pimentel et al. Reference Pimentel, Andow, Dyson-Hudson, Gallahan, Jacobson, Irish, Kroop, Moss, Schreiner, Shepard and Thompson1980). In contrast, organic and low-input systems focus on cultural controls, including mechanical weed control methods such as tillage and cultivation. However, these methods are also environmentally and economically expensive because they can result in soil degradation, require high fossil fuel–derived energy consumption, and increase greenhouse gas emissions (Holland Reference Holland2004). A strategy that is complementary to both conventional and organic methods is to employ an agroecological approach to weed management (Mohler Reference Mohler2001). A key element of such an approach is to utilize crop varieties with enhanced abilities to compete with and tolerate weeds, enabling potential reductions in herbicide application rates (Blackshaw et al. Reference Blackshaw, O’donovan, Harker, Clayton and Stougaard2006; Christensen Reference Christensen1994). Therefore, development of such varieties has been proposed as a fundamental goal in plant breeding programs aiming to maximize productivity and sustainability (Andrew et al. Reference Andrew, Storkey and Sparkes2015; Gallandt and Weiner Reference Gallandt and Weiner2007; Hoad et al. Reference Hoad, Bertholdsson, Neuhoff and Köpke2012).
Crop competitive ability can be separated into two components: 1) the competitive effect or weed-suppressive ability, i.e., the capacity of the crop to reduce weed growth and reproductive success through interference, and 2) the competitive response or weed tolerance; i.e., the ability of the crop to yield despite the presence of weeds (Cahill et al. Reference Cahill, Kembel and Gustafson2005; Lemerle et al. Reference Lemerle, Smith, Verbeek, Koetz, Lockley and Martin2006; Watson et al. Reference Watson, Derksen and Van Acker2009). Weed tolerance may be conferred by suppressive ability if competition involves resource pre-emption at an early stage (Callaway Reference Callaway1992). Alternatively, mechanisms of weed tolerance may be independent of suppressive traits, for example, where there are differential temporal demands for resources with competitors or variations in sensitivities to allelochemicals (Callaway Reference Callaway1992; Goldberg and Landa Reference Goldberg and Landa1991; Jordan Reference Jordan1993). Traits that are consistently expressed under different environmental conditions may have the potential to be applied as heritable selection criteria for further breeding of weed competitive crop varieties (Lemerle et al. Reference Lemerle, Gill, Murphy, Walker, Cousens, Mokhtari, Peltzer, Coleman and Luckett2001).
Variation in the impacts of weeds on crop yield, resulting from differences in crop plant genetics, has been observed in many crops (Callaway Reference Callaway1992). Investigation of the ability of a crop to control weeds through plant competition requires variation in competitive ability among cultivars and identification of weed-suppressive traits, which have been widely documented in other crops including rice (Oryza sativa L.) (de Vida et al. Reference de Vida, Pérez, Emilio, Mackill, Fernández and Fischer2006), wheat (Triticum aestivum L.) (Cosser et al. Reference Cosser, Gooding, Thompson and Froud-Williams1997; Mason and Spaner Reference Mason and Spaner2006; Ogg and Seefeldt Reference Ogg and Seefeldt1999), barley (Hordeum vulgare L.) (Dhima et al. Reference Dhima, Vasilakoglou, Gatsis and Eleftherohorinos2010; Watson et al. Reference Watson, Derksen and Van Acker2009), and soybean [Glycine max (L.) Merr.] (Vollmann et al. Reference Vollmann, Wagentristl and Hartl2010), as well as noncrop species (Gaudet and Keddy Reference Gaudet and Keddy1988; Goldberg and Landa Reference Goldberg and Landa1991). However, few studies have specifically examined the weed-suppressive characteristics of oats, despite their widely accepted superior competitiveness over other cereal crops (Lemerle et al. Reference Lemerle, Verbeek and Coombes1995; Seavers and Wright Reference Seavers and Wright1999; Satorre and Snaydon Reference Satorre and Snaydon1992) and value as a low-input crop in sustainable agricultural systems around the world (Marshall et al. Reference Marshall, Cowan, Edwards, Griffiths, Howarth, Langdon and White2013; Tamm et al. Reference Tamm, Tamm and Ingver2009).
This study examines the ability of husked and naked oat varieties to compete with naturally occurring weed populations under organic conditions. We test three hypotheses: 1) the weed tolerance of winter oat varieties will vary, 2) crop phenotypic traits, such as height, canopy cover, or tillering ability, are important weed-suppressive characteristics in winter oats, and 3) there will be trade-offs between a variety’s competitiveness and its productivity.
Materials and Methods
Study Site
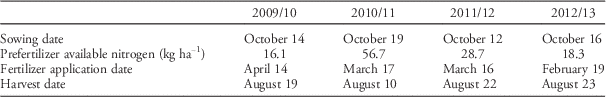
Field trials were conducted with eight winter oat varieties over four years (2009/2010, 2010/2011, 2011/2012, and 2012/2013) at an organically managed (Soil Association certified 1997) site in Suffolk, UK (52°21′37.64′′N, 1°21′28.98′′E). The soil type was a medium-clay loam (clay content 23% to 26% by volume) with slightly alkaline pH (7.4 to 8.0) and moderate soil organic matter levels (23 to 28 g kg−1). Each year’s trials were conducted in a different field, but always in the first cereal position of the rotation, following a 3-year fertility-building grass–clover crop, which was ploughed in the autumn before creating a seed bed. Dominant weed species, with varying abundance across all trial years, included Persian speedwell (Veronica persica Poir.), blackgrass (Alopecurus myosuroides Huds.), annual blue grass (Poa annua L.), creeping bentgrass (Agrostis stolonifera L.), redtop (Agrostis gigantea Roth), quackgrass [Elymus repens (L.) Gould.], broadleaf dock (Rumex obtusifolius L.), curly dock (Rumex crispus L.), and wild mustard (Sinapis arvensis L.). Mechanical weed control was applied in all trial years according to best practice for that year. This generally included cultivations to control weeds in a false seed bed before drilling, as well as tine-weeding and inter-row harrowing throughout the year when appropriate. Dates of field operations and soil fertility each year are shown in Table 1.
Table 1 Sowing dates, soil fertility levels, and fertilizer application and harvest dates for each trial year.
Experimental Design
Separate trials for husked and naked oat varieties were conducted in adjacent areas of the same field each year due to the different harvesting requirements of the two crop types. Five husked oat varieties (‘Balado’, ‘Brochan’, ‘Gerald’, ‘Mascani’, and ‘Tardis’) and three naked oat varieties (‘Bastion’, ‘Mason’, and ‘Racoon’) were tested at two fertility levels (nontreated and 176kgha−1 of organic chicken manure pellets delivering approximately 60kgha−1 of available nitrogen) with three replicates per treatment–variety combination in a randomized complete block design, resulting in a total of 30 plots of husked oat and 18 plots of naked oat per year. Gerald was omitted from year one due to lack of available seed. Plots measuring 1.2 by 10.2 m were sown using a six-coulter plot drill and 20-cm row spacing. Target plant populations were set according to rates commonly used in organic systems of 425 plants m−2 (Lampkin and Padel Reference Lampkin and Padel2004).
Crop Assessments
Plant emergence and plant establishment were assessed by counting plant numbers in two randomly placed 0.25-m−2 quadrats per plot when the crop was at growth stage (GS) 10 to 12 (Zadoks et al. Reference Zadoks, Chang and Konzak1974) and GS 25, respectively. Total percent ground cover of all weed species combined was assessed for early-season weed cover when the crop entered stem elongation (GS 31) and for post-harvest weed cover immediately after harvest, in two 0.25-m−2 quadrats per plot; small markers were used to enable the same area to be assessed at both timings. Crop leaf area index (LAI) was measured using a Sun Scan Canopy Analysis System type SS1 (Delta-T Devices, Cambridge, UK), at three time points during the year: early-season (stem elongation, GS 31), mid-season (booting, GS 41), and late-season(mid flowering, GS 65). Crop height to the base of the panicle was assessed at flowering (GS 60) for ten randomly selected stems per plot. Stem density was assessed after flowering (GS 69) by counting the number of fertile tillers along two randomly selected 1-m row lengths per plot. Foliar diseases including crown rust (Puccinia coronata f. sp.), powdery mildew (Blumeria graminis f. sp. avenae), and leaf spot (Pyrenophora avenae) were assessed after flowering (GS 65) and before leaf senescence by estimating the percent cover of each disease symptom on ten flag leaves per plot (James Reference James1971). Prior to harvest, plants were destructively sampled to ground level over a 1-m row length twice per plot to determine grain numbers, grain weight per stem, and straw biomass. Total harvested plot grain yield (kg ha−1) (adjusted to 15% humidity) was recorded at harvest.
Statistical Analysis
As detailed above, the field trials for husked and naked oats were independent of one another and were therefore treated separately for all analyses. ANOVA with year as the single factor was used to determine the differences in average weed pressures among the four trial years. Analysis of covariance (ANCOVA) was used to determine the most influential factors affecting grain yield and varietal differences in weed tolerance. Variety and fertility level were included as factors, and early-season percent weed cover and disease scores were included as covariates. Year was not included as a factor in this analysis because weed and disease levels were linked to year and therefore confounding. Data for early-season weed cover were log e (x+1) transformed to normalize the distribution of residuals (confirmed using the Shapiro-Wilk test). Other studies calculated weed tolerance as yield loss in weedy compared to weed-free treatments (Callaway Reference Callaway1992). In the current study, we calculated weed tolerance in a similar way for each crop variety, as the estimated yield loss between 0% and 1.72% early-season weed cover regressed over four trial years. This measure of weed tolerance was used because it gives a value of 1 after loge (x+1) transformation. Varietal differences in weed-suppressive traits and percent early-season and post-harvest weed cover were identified using ANOVA with variety, fertilizer treatment, and year as interacting fixed factors, and block nested within year as a random factor. Values of crop height and mid-season LAI were log e transformed to ensure normal distribution of model residuals. ANOVA and ANCOVA models were simplified from full models including all possible two-way interactions by stepwise reduction based on Akaike’s information criterion and pairwise comparisons between varieties were made using Tukey’s HSD test. All statistical analyses were conducted using R version 3.0.2 (R Core Team 2013).
Path Analysis
Path analysis was used to identify weed-suppressive traits and trade-offs between competitiveness and productivity. Path analysis is a statistical tool to evaluate the implied causative effects of confounding variables within a hypothesized theoretical model (Land Reference Land1969). Standardized path coefficients were generated across multiple regression and correlation models. Eight sequential hypothesized partial regression models were created for each crop and weed path analysis for husked and naked oats. The first included establishment as a dependent variable with emergence as a single independent variable. The second included early-season LAI as a dependent variable with establishment as a single independent variable. The third included mid-season LAI as a dependent variable with early-season LAI and height as independent variables. The fourth included late-season LAI as a dependent variable with mid-season LAI and height as independent variables. The fifth included stem density as a dependent variable with establishment as a single independent variable. The sixth included early-season weed cover as a dependent variable with establishment and early-season LAI as independent variables. The seventh included post-harvest weed cover as a dependent variable with early-season weed cover, early-season LAI, mid-season LAI, late-season LAI, stem density, and height as independent variables. The eighth included grain yield as a dependent variable with thousand grain weight, grains per stem, stem density, height, post-harvest weed cover, early-season weed cover, late-season LAI, and mid-season LAI as independent variables. Correlations were calculated between yield components (thousand grain weight, stem density, and grains per stem), because these relationships were considered noncausative. Indirect effects between one independent variable and the dependent variable via a second independent variable were calculated by multiplying the direct path of the two independent variables with the direct path from the second variable to the dependent variable (Pantone et al. Reference Pantone, Baker and Jordan1992). R package lavaan was used to carry out path analysis (Rosseel Reference Rosseel2012). Path analyses for husked and naked oats were calculated across all four years. Only significant effects (P<0.05) were included in path models.
Broad Sense Heritability
Broad sense heritability (H 2) of suppressive traits was estimated from the ANOVA using the following formula:
 $$H^{{\rm 2}} =\rsigma ^{2} _{{\rm G}} \left[ {\rsigma ^{2} _{{\rm G}} {\plus}{\rm }\left( {\rsigma ^{2} _{{{\rm GE}}} /E} \right){\plus}\left( {\rsigma ^{2} _{{\rm e}} /r} \right)} \right]$$
$$H^{{\rm 2}} =\rsigma ^{2} _{{\rm G}} \left[ {\rsigma ^{2} _{{\rm G}} {\plus}{\rm }\left( {\rsigma ^{2} _{{{\rm GE}}} /E} \right){\plus}\left( {\rsigma ^{2} _{{\rm e}} /r} \right)} \right]$$
where σ2 G is the genetic variance, σ2 GE is the genotype ×experiment interaction variance, σ2 e is the residual variance, E is the number of experiments, and r is the number of replicates per crop variety (Nyquist and Baker Reference Nyquist and Baker1991).
Results and Discussion
This study investigated the capacity of winter oat varieties to yield despite competition from weeds. Percent weed ground cover varied significantly over the four years for both husked (Table 2, F3=11.07, P<0.001) and naked oats (Table 2, F3=3.84, P=0.013), and was found to have the greatest impact on yield compared to other factors. After accounting for effects of crop variety on yields in husked (Table 3, F4,17=11.65, P<0.001) and naked (Table 3, F2,9=15.47, P<0.001) oats, early-season weed cover had the largest effect on reducing crop yield of both husked (Table 3, F1,17=72.85, P<0.001) and naked (Table 3, F1,9=14.76, P<0.001) oats. Average crop yields were approximately 16% lower for husked and 25% lower for naked oats in the year with the greatest early-season weed cover (2011) compared to the year with the least early-season weed cover (2009). Total disease on the flag leaf was a significant yet less influential covariate in husked oats (Table 3, F1,17=22.94, P<0.001) but was nonsignificant in naked oats. Added fertilizer did not have a significant effect on yield in husked or naked oats, indicating that nutrient levels in the field were ample following the three-year grass–clover crop. However, added fertilizer decreased post-harvest weed cover in some trial years in husked and naked oats (Table 4). This suggests that addition of fertilizer affects crop–weed competitive interactions in favor of the oat crop (Blackshaw and Brandt Reference Blackshaw and Brandt2008) and highlights oats as a generally competitive and resource use efficient crop.
Table 2 Differences in mean early-season weed cover among trial harvest years in husked and naked oat trials; n=48. Values with the same letter within husked and naked oats do not differ significantly by Tukey’s HSD test (P>0.05).

Table 3 ANCOVA of grain yield in husked and naked oat experiments over four years. Results include each factor or covariate separately, as well as two-way interactions between them. Models were simplified by stepwise reduction based on Akaike’s information criterion.

a Abbreviations: DF, degrees of freedom; MS, mean squares.
Table 4 Effects of added fertilizer on post-harvest weed cover in husked and naked oat trials over all trial years and each year individually. N=15 for husked oats and N=9 for naked oats. Asterisks indicate significant effects: *, P<0.05; **, P<0.01.

Weed Tolerance
Varietal differences in weed tolerance were found. Significant interactions between crop variety and early-season weed cover effected yield of husked (Table 3, F4,17=7.87, P<0.001) but not naked oats, indicating differences in varietal yield response to weed competition among the husked oat varieties only. The slope of each line in Figure 1 indicates the weed tolerance of each crop variety, which is given in Table 5 along with a summary of suppressive traits for husked and naked oat varieties. For example, Balado, the semidwarf variety with the poorest weed tolerance, was predicted to suffer a 35% yield loss due to weeds in the year that weed pressure was greatest (2012) (Figure 1a). On the other hand, Racoon, the tallest variety, had the greatest weed tolerance among naked varieties (Figure 1b). This suggests that height was an important trait for weed tolerance, whereas other competitive traits such as crop tillering and LAI were not found to be important. High tillering varieties, such as Bastion, and varieties with high LAI, such as Brochan, had only moderate weed tolerance (Table 5, Figure 1). The importance of height for weed tolerance is also supported by studies in wheat, which suggest that maintaining a height advantage is an effective strategy for increasing weed tolerance because it allows the crop to escape the competitive shading effect of weeds (Blackshaw Reference Blackshaw1994; Cousens et al. Reference Cousens, Barnett and Barry2003; Moss Reference Moss1985). This may be less applicable in the case of climbing or taller-growing weed species such as catchweed bedstraw (Galium aparine L.) or wild oat (Avena fatua L.) (Cousens et al. Reference Cousens, Barnett and Barry2003).

Figure 1 Relationship between early-season weed cover and grain yield in (a) husked and (b) naked oat varieties. The slope of each line indicates weed tolerance for each variety. N=24 for all varieties except Gerald, for which n=18. An overall significant effect of early-season weed cover yield was found across all varieties in husked and naked oats (P<0.001). Asterisks indicate significance levels of this relationship per variety: *, P<0.05; **, P<0.01; ***, P<0.001. The weed tolerance of Balado was significantly less than that of all other husked varieties, and the other varieties did not significantly differ from each other (a). No significant differences in weed tolerance were found among naked oat varieties (b).
Table 5 Values of weed tolerance and mean values of weed-suppressive traits for each crop variety. Values of weed tolerance represent estimated t ha−1 yield loss between 0% and 1.72% weed cover. Estimates of broad sense heritability (H 2) indicate how consistently varietal differences are expressed in different years. N=24 for all varieties except Gerald, for which N=18. Abbreviation: LAI, leaf area index.

Values followed by the same letter within husked or naked oats do not differ significantly by Tukey’s HSD test (P>0.05).
Weed Suppression
The relative importance of specific weed-suppressive traits was assessed using path analysis. Early-season weed cover was influenced by crop establishment (Figure 2, husked oats direct path −0.27, P<0.01; Figure 3, naked oats direct path −0.30, P < 0.05). The early-season percent mean weed cover was 21% lower in the crop variety with the highest (Mascani) compared to the lowest (Balado) mean establishment rates (Figure 4a). Variation in early-season LAI did not affect early-season weed cover (%), indicating that aboveground competitive interactions were not effective at this stage of crop establishment. Crop emergence rates also had an indirect negative effect on early-season weed cover through their relationship with crop establishment in both the husked and naked oat trials (−0.11 and −0.21, respectively). In the husked oat trials, crop establishment indirectly affected post-harvest weed cover via both stem density and early-season weed cover (total indirect path −0.19). This comparison cannot be made with the naked oat trials as there was no significant path via stem density. Post-harvest weed cover was also directly affected by early-season weed cover in both husked and naked oat trials (direct paths 0.38, P<0.001 and 0.46, P<0.001, respectively) (Figures 2 and 3). These results highlight the importance of competitive interactions early in the year during crop establishment and their consequent effects later in the year. Rapid establishment and growth rates have also been identified as important competitive traits in crops (Cosser et al. Reference Cosser, Gooding, Thompson and Froud-Williams1997; de Vida et al. Reference de Vida, Pérez, Emilio, Mackill, Fernández and Fischer2006; Lemerle et al. Reference Lemerle, Verbeek, Cousens and Coombes1996) and noncrop plant species (Gaudet and Keddy Reference Gaudet and Keddy1988; Goldberg and Landa Reference Goldberg and Landa1991). Mid-season LAI had a negative influence on post-harvest weed cover in the husked oat trial (direct paths −0.40, P<0.05) (Figure 2). Mean post-harvest weed cover was 41% lower in the crop variety with the highest (Brochan) compared to that with the lowest (Gerald) mid-season LAI (Figure 4b). This significant effect of mid-season LAI, rather than early- or late-season LAI, suggests that this stage in the crop’s development before panicle emergence (GS 41) is the most critical period for weed suppression, probably due to competition for light. Stem density had the greatest negative influence on post-harvest weed cover compared to other suppressive traits such as height or LAI, in both the husked and naked oat trials (direct path −0.44, P<0.001 and −0.38, P<0.001) (Figures 2 and 3). This supports evidence that cereal varieties with high tillering ability can effectively suppress weeds (Hoad et al. Reference Hoad, Bertholdsson, Neuhoff and Köpke2012; Lemerle et al. Reference Lemerle, Verbeek, Cousens and Coombes1996). Crop height is widely considered to be a valuable competitive trait for light competition (Didon and Hansson Reference Didon and Hansson2002; Drews et al. Reference Drews, Neuhoff and Köpke2009; Gooding et al. Reference Gooding, Thompson and Davies1993). However, it was not found to have a direct suppressive effect on post-harvest weed cover in the current study. Height did have an indirect effect on post-harvest weeds (indirect path 0.20) via mid-season LAI in the husked oat trial, yet greater height did not always confer a high LAI. For example, Gerald was significantly taller than Brochan, but had a significantly lower mid-season LAI, which is likely due to Gerald’s particularly upright, erectophile leaf angle. Previous studies in wheat have also suggested that morphological traits such as planophile leaf angle and growth habit contribute more to LAI than height alone (Seavers and Wright Reference Seavers and Wright1999).

Figure 2 Path diagram showing the hypothesized direct effects of competitive traits on husked oat grain yield and weed cover. Pertinent variables include establishment rates, tillering ability (stem density), early-season leaf area index (LAI), and straw height. TGW, thousand grain weight. Black arrows indicate positive effects and grey arrows indicate negative effects. Only standardized direct path coefficients (unidirectional arrows) and correlation coefficients (double-headed arrows) significant at P<0.05 are included in the diagram. Asterisks indicate significance levels: *, P<0.05; **, P<0.01; ***, P<0.001.

Figure 3 Path diagram showing the hypothesized direct effects of competitive traits on naked oat grain yield and weed cover. Pertinent variables include establishment rates, tillering ability (stem density), early-season leaf area index (LAI), and straw height. TGW, thousand grain weight. Black arrows indicate positive effects and grey arrows indicate negative effects. Only standardized direct path coefficients (unidirectional arrows) and correlation coefficients (double-headed arrows) significant at P<0.05 are included in the diagram. Asterisks indicate significance levels: *, P<0.05; **, P<0.01; ***, P<0.001.

Figure 4 Mean values of (a) early-season weed cover and (b) post-harvest weed cover among husked and naked oat varieties over four trial years. N=24 for all varieties except Gerald, for which N=18. Values with the same letter within husked and naked oats do not differ significantly by Tukey’s HSD test (P>0.05).
Varieties with Superior Weed Suppressive Ability
To investigate whether certain varieties have greater weed suppression ability than others, differences in traits identified as beneficial for weed suppression were compared among husked and naked oat varieties. Suppressive traits that differed significantly among crop varieties are outlined in Table 5. To be a useful selection criterion, suppressive traits must have a high heritability; being consistently expressed over different environments. Values of broad sense heritability in Table 5 indicate the proportion of the genetic to total variance in a phenotype; thus, how consistent varietal differences in traits are between years. Although LAI was identified as a useful suppressive trait, it had relatively low heritability in this study. It could be that there was unavoidable error when measuring genotypic differences in LAI, perhaps caused by the additional shading effect of weeds as well as the crop or inconsistent crop establishment rates each year. Plant height, which is a much more heritable trait and is linked to LAI, could be a useful indirect selection criterion for LAI when selection is not in a target environment such as low-input systems. Although stem density had a high heritability, it may also be strongly influenced by crop establishment rates early in the year. However, varietal differences in crop establishment rates are more year-dependent with low heritability, possibly due to other factors such as varying seed quality among varieties, whilst tillering ability is a more heritable trait despite this effect (Table 5). Combinations of environmental factors influence tillering in cereals, including light quality, temperature, and water and nutrient availability (Laude Reference Laude1972). Therefore, a crop variety’s ability to tiller may be indicative of its ability to use and compete effectively for mainly below-ground resources early in the year. The current study highlights the potential to select for tillering ability and tiller survival when under high weed and crop density, to ensure effective weed suppression. Furthermore, there may be potential to optimize these traits for enhanced performance at higher seeding rates (Weiner et al. Reference Weiner, Griepentrog and Kristensen2001). Selection of breeding material in competitive environments, under high weed levels and crop density, would enable better selection of environment-dependent traits relating to weed tolerance and suppression (Lemerle et al. Reference Lemerle, Verbeek, Cousens and Coombes1996). Further research is needed to examine competitive interactions in a wider range of environments, such as different sites or different positions in organic rotation where soil fertility is lower and weed pressure may be higher.
Crop Yield and Grain Quality
Potential trade-offs between competitiveness and productivity must be considered in any breeding program aiming to develop cultivars with high competitive ability (Vandeleur and Gill Reference Vandeleur and Gill2004). Weed-suppressive traits, such as LAI, can increase intracrop competition for light and allocation of resources to vegetative growth rather than reproduction. In the absence of significant weed competition, this may limit the potential productivity and grain yield of the crop (Weiner Reference Weiner1988). Potential yield in a monoculture is often reduced by the increased competitive ability of individuals (Creissen et al. Reference Creissen, Jorgensen and Brown2013; Donald Reference Donald1968; Hamblin and Rowell Reference Hamblin and Rowell1975). However, weed-suppressive traits identified in path analysis did not trade-off against grain yield in this study. Mid-season LAI, which reduced post-harvest weed cover in the husked oat trial, did not have a negative impact on grain yield. Straw height, which had an indirect suppressive effect on weeds, had a positive influence on grain yield in both the husked and naked oat varieties (direct paths=0.23, P=0.01 and 0.25, P=0.008, respectively) (Figures 2 and 3). Crop height was also associated with weed tolerance and so is a key trait related to yielding ability when weed competition is sufficiently high. Stem density had the most suppressive effect on weeds and also had a positive effect on grain yield in both husked and naked oat trials (direct paths=0.33, P<0.01 and 0.58, P<0.001, respectively) (Figures 2 and 3). It may be that the effect of weed pressure in this study sufficiently masked any effect of intraspecific crop competition. The contrary is likely to be observed in weed-free conventional farming systems. A strategy for increasing competitive ability while not limiting yield potential in the absence of weeds may focus on enhancing competitive traits such as LAI and vegetative growth early in the year, while limiting competitive traits later in the year, when resources are allocated to grain filling (de Vida et al. Reference de Vida, Pérez, Emilio, Mackill, Fernández and Fischer2006; Dingkuhn et al. Reference Dingkuhn, Johnson, Sow and Audebert1999).
In addition to using cultivars with high tillering ability, increasing sowing rate can more directly increase weed suppression (Korres and Froud-Williams Reference Korres and Froud‐Williams2002). However, the potential yield benefit at higher seed rates may be negated by a reduced harvest index and an increase in the potential lodging risk at high plant densities (Mohler et al. Reference Mohler2001). It may also result in poorer oat grain quality with lower specific weight, increased screenings in the milling process, and aborted grains as a result of greater competition for photosynthates between panicles (Browne et al. Reference Browne, White and Burke2003, Reference Browne, White and Burke2006). This is supported by a compensatory effect between the yield components in this study. In both the husked and naked oat trials, strong and consistent negative correlations between stem density and grains per stem (correlation coefficient=−0.75, P<0.001 and −0.77, P<0.001, respectively) (Figure 2 and 3), and stem density and thousand grain weight (correlation coefficient=−0.36, P<0.001 and −0.62, P<0.001, respectively) (Figures 2 and 3), were observed.
Mechanisms of crop–weed competitive interactions discussed here offer insights into an underutilized cultural weed management approach that can reduce herbicide use in conventional systems and soil cultivations in organic farming systems. Problems with herbicide resistance and soil degradation highlight the fact that no single approach to weed control remains effective in the long term (Heap Reference Heap2014). However, cereal breeding for conventional agriculture intended to increase yield and reduce lodging risk with high fertilizer applications by introducing dwarfing genes to increase harvest index (Peng et al. Reference Peng, Richards, Hartley, Murphy, Devos, Flintham, Beales, Fish, Worland, Pelica and Sudhakar1999) has been counterproductive for competitive ability and crop resilience in low-input or organic systems (Lammerts van Bueren et al. Reference Lammerts van Bueren, Jones, Tamm, Murphy, Myfers, Leifert and Messmer2011). In summary, this study highlights the potential for enhancing competitive interactions between crop and weeds through weed suppression, and maintenance of yield despite competition from weeds. Suppressive traits, including crop establishment early in the year, tillering ability, and LAI, were found to enhance competitive ability by ensuring effective use of, and pre-emptive competition for, light during the critical weed growth period in the spring. Tolerance to weed competition was conferred by crop height facilitating the maintenance of a competitive advantage over weeds. In general, an effective strategy may be a combination of both weed suppression and weed tolerance, the first to reduce weed growth and seed production and seedbanks (Cosser et al. Reference Cosser, Gooding, Thompson and Froud-Williams1997) for long-term management of weed population dynamics, and the second to maintain yield in environments, such as organic farms, where weed competition is unavoidable. In conclusion, the use of competitive oat varieties is integral to sustainable weed management strategies aimed at reducing dependence on unsustainable and potentially environmentally damaging herbicides and cultivation.
Acknowledgements
The QUOATS project, led by Aberystwyth University, was funded by the Agriculture and Horticulture Development Board (AHDB) and industry partners, and was jointly sponsored by the Biotechnology and Biological Sciences Research Council (BBSRC), Defra through the Sustainable Arable LINK Programme, European Regional Development Funding through the Welsh Assembly Government’s Academic Expertise for Business (A4B) Programme, and the Scottish Government Contract Research fund. The authors thank Louisa Winkler and Thomas Hughes, who contributed to field trials, as well as Martin Wolfe and Paul and Mark Ward for excellent management of the trial site and agronomic advice.










 Open access
Open access