



Introduction
The survival of the bear cuscus Ailurops ursinus, a phalangerid marsupial endemic to Sulawesi and its surrounding islands, is threatened by poaching for consumption and illegal trade (Shepherd et al., Reference Shepherd, Emerson, Janssen and Morgan2018; Bailey et al., Reference Bailey, Hilser, Siwi, Lawe, Waterman and Loffeld2022), deforestation (Salas et al., Reference Salas, Dickman, Helgen and Flannery2019; Repi et al., Reference Repi, Masyud, Mustari and Prasetyo2020) and habitat loss and degradation (Nugraha et al., Reference Nugraha, Andriyani and Marliana2021). The species inhabits tropical lowland forest (Dwiyahreni et al., Reference Dwiyahreni, Kinnaird, O'Brien, Supriatna and Andayani1999; Nugraha & Mustari, Reference Nugraha and Mustari2017; Salas et al., Reference Salas, Dickman, Helgen and Flannery2019) across a small number of ecosystem types (limestone, non-dipterocarp, monsoon deciduous and monsoon evergreen forests), with occasional occurrence in shrubland, farmland and mixed farmland (Martin et al., Reference Martin, Monkhouse, O'Connell, Analuddin, Karya and Priston2019; Nugraha et al., Reference Nugraha, Andriyani and Marliana2021), and is heavily reliant on continuous canopy cover for dispersal. Forest fragmentation has hindered the species’ movement and limited access to food and shelter (Nugraha et al., Reference Nugraha, Andriyani and Marliana2021). The bear cuscus is believed to be the only diurnal cuscus species, and because of its slow movement and large size (Nowak, Reference Nowak2018; Salas et al., Reference Salas, Dickman, Helgen and Flannery2019; Mustari, Reference Mustari2020) it can be easily captured by people (Nugraha & Mustari, Reference Nugraha and Mustari2017). Given the absence of large mammalian carnivores in Sulawesi forests, people are the species' main predator (Rejeki, Reference Rejeki2018).
As the largest landmass in Wallacea, Sulawesi is one of the greatest biodiversity hotspots (Struebig et al., Reference Struebig, Aninta, Beger, Bani, Barus and Brace2022) and is notable for its high rate of endemism, with endemic species continuing to be discovered (Esselstyn et al., Reference Esselstyn, Achmadi, Handika, Swanson, Giarla and Rowe2021). It is also the westernmost extent of marsupials in Asia and is home to four endemic phalangerids, two of which are categorized as threatened on the IUCN Red List: the Vulnerable bear cuscus and Critically Endangered Talaud bear cuscus Ailurops melanotis. The Sulawesi dwarf cuscus Strigocuscus celebensis is categorized as Near Threatened and the Peleng cuscus Strigocuscus pelengensis as Least Concern (Farida, Reference Farida, Ubaidillah and Abinawanto2022). Despite its status and declining population trend (Salas et al., Reference Salas, Dickman, Helgen and Flannery2019), the bear cuscus is not protected by Indonesian law (Shepherd et al., Reference Shepherd, Emerson, Janssen and Morgan2018; Farida, Reference Farida, Ubaidillah and Abinawanto2022) nor listed in the CITES appendices, even though it is commonly traded in Sulawesi and abroad (Rejeki, Reference Rejeki2018; Shepherd et al., Reference Shepherd, Emerson, Janssen and Morgan2018; Latinne et al., Reference Latinne, Saputro, Kalengkongan, Kowel, Gaghiwu and Ransaleleh2020). It inhabits several large protected areas of > 200,000 ha, including Bogani Nani Wartabone and Lore Lindu National Parks, and Morowali, Tangkoko Batuangus and Gunung Dua Saudara Nature Reserves, as well as various other forest reserves (Salas et al., Reference Salas, Dickman, Helgen and Flannery2019). All of these protected areas are located in the provinces of North and Central Sulawesi, thus these bear cuscus populations are separated from those in South Sulawesi. The bear cuscus was previously protected by Indonesian law during 1999–2018, when it was classified as Phalanger ursinus, but no longer has any legal protection.
Research on the bear cuscus is limited and tends to focus on characteristics such as the species’ morphology (Klećkowska-Nawrot et al., Reference Klećkowska-Nawrot, Goździewska-Harłajczuk, Darska, Barszcz and Janeczek2019; Goździewska-Harłajczuk et al., Reference Goździewska-Harłajczuk, Klećkowska-Nawrot, Hamouzová and Čížek2020), acoustic behaviour (Bool et al., Reference Bool, Slaymaker, Magrath, Arfian, Karya, Analuddin and Courtney Jones2020), diet (Dwiyahreni et al., Reference Dwiyahreni, Kinnaird, O'Brien, Supriatna and Andayani1999; Achmad et al., Reference Achmad, Ngakan, Maulany and Asrianny2016b; Farida & Dahruddin, Reference Farida and Dahruddin2008; Nugraha & Mustari, Reference Nugraha and Mustari2017; Nugraha et al., Reference Nugraha, Andriyani and Marliana2021), daily movements (Achmad et al., Reference Achmad, Ngakan, Maulany and Asrianny2016a) and behaviour (Talumepa et al., Reference Talumepa, Wungow, Poli and Rimbing2016; Nugraha & Mustari, Reference Nugraha and Mustari2017; Sara et al., Reference Sara, Bachtiar and Puspaningrum2020). Reports on bear cuscus ecology are comparatively scarce, although research has been published on the characteristics of its habitat (Nugraha & Mustari, Reference Nugraha and Mustari2017; Khairunnisa & Mustari, Reference Khairunnisa and Mustari2018), its habitat use (Nugraha et al., Reference Nugraha, Andriyani and Marliana2021) and its distribution (Martin et al., Reference Martin, Monkhouse, O'Connell, Analuddin, Karya and Priston2019).
Nothing is known about the species' potential distribution or the availability of suitable habitat, which are important factors in determining areas of conservation priority. The bear cuscus also has the potential to serve as a so-called Cinderella species, a term coined by Smith et al. (Reference Smith, Veríssimo, Isaac and Jones2012) to describe an overlooked potential flagship species. Our study therefore aimed to predict the habitat suitability and potential distribution of the bear cuscus in South Sulawesi using maximum entropy (MaxEnt) modelling, and to subsequently suggest priority areas for its conservation, taking into account land-use type and anthropogenic threats. To our knowledge, this is the first study of its kind undertaken for the bear cuscus, and, given the dearth of information on the species, it can provide a more complete picture of its distribution and guide management of its population and habitat.
Study area
We collected our data in the protected areas of Bantimurung Bulusaraung National Park and Hasanuddin University Educational Forest in South Sulawesi, Indonesia (Fig. 1). Both areas are key habitats for the bear cuscus and other endemics such as the moor macaque Macaca maura (Riley et al., Reference Riley, Albani, Zak, Germani, Rothman, Carosi and Ngakan2023) and Makassar tarsier Tarsius fuscus (Andriyani et al., Reference Andriyani, Nugraha and Marliana2021). Unabated deforestation, both inside and adjacent to the protected areas and stemming primarily from the expansion of agriculture, mining and urban development, has steadily degraded and fragmented these habitats (Mustari et al., Reference Mustari, Surono and Mansyur2011; Broto & Setiawan, Reference Broto and Setiawan2013; Dwiyahreni et al., Reference Dwiyahreni, Fuad, Sunaryo, Soesilo, Margules and Supriatna2021). Unprotected buffer areas adjacent to the National Park and Educational Forest serve as a natural corridor for the species’ dispersal (Nugraha, Reference Nugraha2021).
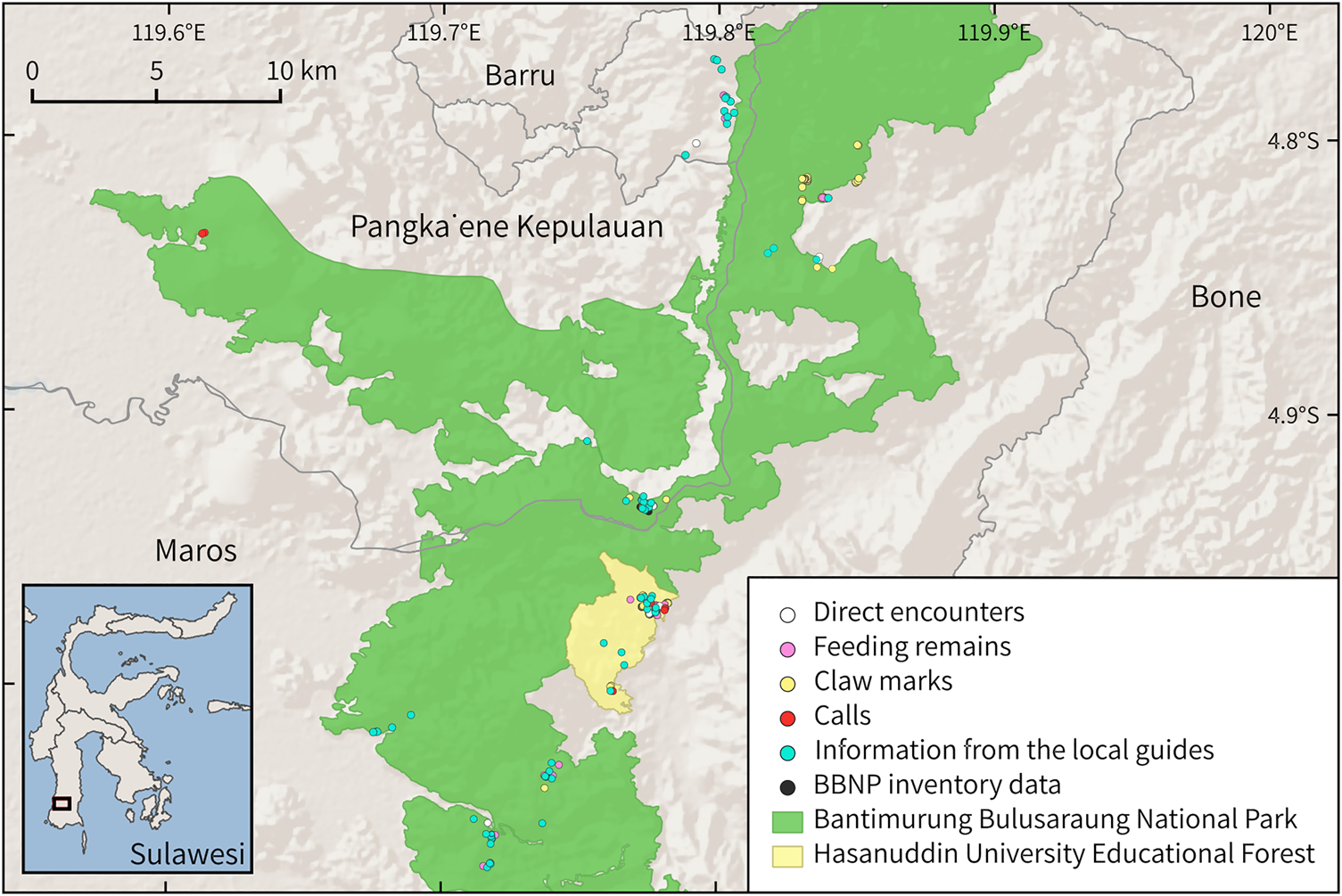
Fig. 1 Bear cuscus Ailurops ursinus records in Bantimurung Bulusaraung National Park (BBNP) and Hasanuddin University Educational Forest, South Sulawesi, Indonesia. All information reported by local guides relates to direct sightings of the species. (Readers of the printed journal are referred to the online article for a colour version of this figure.)
Methods
Model area coverage and variables
To increase the accuracy of our habitat suitability model, we limited the area of coverage to southern South Sulawesi based on land-cover type; the northern part of the modelling area is considered unsuitable habitat for the bear cuscus as it is mostly dominated by rice fields, other agriculture and settlements, essentially preventing northward movement of the marsupial (Nugraha, Reference Nugraha2021).
We collected occurrence data during October 2020–January 2021 by recording any direct observations of the species, indirect signs (i.e. claw marks, feeding remains and calls; Supplementary Plates 1 & 2) and reports of direct sightings provided by local guides and residents, as well as using inventory data from a survey of the bear cuscus carried out in the National Park in 2019 (Fig. 1). We searched for 60 days between 07.00–11.00 and 13.00–18.00, walking 45 tracks of various lengths on and off footpaths, totalling 240.34 km (Supplementary Fig. 1). Because the longest recorded daily distance traversed by a bear cuscus is 78 m (Achmad et al., Reference Achmad, Ngakan, Maulany and Asrianny2016a), to minimize spatial autocorrelation we used a threshold of 90 m to eliminate encounter points using the species thinning technique in the package spThin (Aiello-Lammens et al., Reference Aiello-Lammens, Boria, Radosavljevic, Vilela and Anderson2015; Condro et al., Reference Condro, Prasetyo, Rushayati, Santikayasa and Iskandar2021) in R 3.6.1 (R Core Team, 2019), leaving 103 data points for use in the model construction.
We included eight environmental variables in the model (Supplementary Table 1): (1) elevation, because the species is known to inhabit ecosystems of various elevations (Nugraha et al., Reference Nugraha, Andriyani and Marliana2021); (2) slope, as steeper areas tend to be less accessible to predators (including people) and thus potentially safer for and preferred by arboreal mammals (Kusumanegara et al., Reference Kusumanegara, Kartono and Prasetyo2017; Iwanda et al., Reference Iwanda, Prasetyo and Rinaldi2019); (3) land surface temperature, a climatic factor that can explain the presence of a species in an area (Lindenmayer et al., Reference Lindenmayer, Nix, McMahon, Hutchinson and Tanton1991); (4) normalized difference vegetation index (NDVI), which serves as a proxy for vegetation cover (Suárez-Seoane et al., Reference Suárez-Seoane, Osborne and Rosema2004), indicating food availability and potential habitat (Youngentob et al., Reference Youngentob, Yoon, Stein, Lindenmayer and Held2015); distance from the nearest path (5) and settlement (6), as anthropogenic disturbances apply pressure on wildlife and increase the chances of being hunted by people (Dewi et al., Reference Dewi, Prasetyo and Rinalfi2007); (7) distance from the nearest river, as water sources are important for the survival of most wildlife species (Sodik et al., Reference Sodik, Pudyatmoko, Yuwono, Tafrichan and Imron2020); and (8) land-cover type, which refers to the layers of soils, biomass and human structures that cover the land surface (Verburg et al., Reference Verburg, Neumann and Nol2011), and is closely related to food sources and habitat conditions (Nugraha, Reference Nugraha2019). In our study, land cover comprised 18 categories: primary forest, secondary forest, plantation forest, shrubland, plantation, settlement, open land, savannah, water body, secondary mangrove forest, thicket swamp, farmland, mixed farmland, rice field, pond, airport/seaport, mining area and swamp.
We created elevation, slope, NDVI and land surface temperature maps in Google Earth Engine (Google, USA), based on Rijal (Reference Rijal2020), with the following modification: we retrieved land surface temperature data from the Landsat 8 thermal band by deriving the values from NDVI and the top of the atmosphere spectral radiance. We used QGIS 3.18.1 (QGIS, 2021) to create land-cover maps based on information from the Indonesian Ministry of Environment and Forestry, and to measure distances to the nearest path, settlement and river. The types of path data we used in model construction were patrol (footpaths), local (rural roads) and arterial roads (highways). We masked the maps according to the model area of coverage at a 30 m pixel resolution projected to the WGS 84/UTM zone 50S coordinate system and converted to the ASCII format.
Model construction and evaluation
We performed a multicollinearity test in R using random points from the extracted values of each environmental variable as the sample, followed by a stepwise regression to obtain the variance inflation factor value (Sodik et al., Reference Sodik, Pudyatmoko, Yuwono, Tafrichan and Imron2020; Widyastuti et al., Reference Widyastuti, Perwitasari-Farajallah, Prasetyo, Iskandar and Setiawan2020). We only used variables with a variance inflation factor value of < 10 to build the model (Dormann et al., Reference Dormann, Elith, Bacher, Buchmann, Carl and Carré2013; Widyastuti et al., Reference Widyastuti, Perwitasari-Farajallah, Prasetyo, Iskandar and Setiawan2020).
We used the MaxEnt algorithm in MaxEnt 3.4.4 to build the habitat model (Phillips et al., Reference Phillips, Anderson and Schapire2006; Elith et al., Reference Elith, Phillips, Hastie, Dudík, Chee and Yates2011), with 75% of the bear cuscus presence points serving as the training data (77 occurrences) and 25% serving as the test data (26 occurrences). We used 10 replicates with the subsample run type to create the model (Nugraha, Reference Nugraha2021). We evaluated the performance of the model based on the mean value of the area under the receiver operating characteristic curve (AUC; Fielding & Bell, Reference Fielding and Bell1997), where the model accuracy was considered to be low, moderate or high at values of 0.5–0.699, 0.7–0.899 or ≥ 0.9, respectively (Manel et al., Reference Manel, Williams and Ormerod2001). We also performed a jackknife test to assess the contribution of each environmental variable to the model (Elith et al., Reference Elith, Phillips, Hastie, Dudík, Chee and Yates2011).
Potential spatial distribution and priority areas for bear cuscus conservation
We used a 10th percentile training presence as the threshold to determine habitat suitability for the bear cuscus, resulting in a mean pixel value of 0.1070. We then categorized pixels into five levels of suitability: unsuitable (pixel value < 0.1070), marginally suitable (0.1070–0.3303), moderately suitable (0.3304–0.5535), mostly suitable (0.5536–0.7768) and highly suitable (> 0.7768). We then overlaid the habitat suitability map with the forest areas according to their government-assigned function (i.e. conservation forest, protection forest or production forest) to predict the spatial distribution of the bear cuscus within each. These government-assigned forest functions were based on Law No. 5 of 1990 and No. 41 of 1999, wherein the main function of a conservation forest (which includes national parks and nature reserves) is to maintain biodiversity and preserve existing ecosystems, with reserves and national parks both limiting anthropogenic activities to research and education, but with the latter also allowing utilization of natural resources and tourist activities. A protection forest has primarily protective and life-supporting functions for humans, such as regulating water, preventing flooding or erosion, or maintaining soil fertility, with selective allowances for non-forestry usages and construction. A production forest is designated for the production of both timber and non-timber commodities, with logging and open-cast mining permitted under specific provisions. Conservation forests are preservatory, and forest rehabilitation is allowed only in the other forest types. Species should thus face the least disturbance in conservation forests and the most disturbance in production forests. With this in mind, we identified bear cuscus conservation regions in southern South Sulawesi based on the dominating designated forest function. We also subsequently overlaid the bear cuscus habitat suitability map with the active legal mining areas in South Sulawesi to determine the overlap of potential bear cuscus distribution within these sites.
Results
We recorded 209 bear cuscus encounters, comprising 16 direct sightings and 193 indirect signs (25 feeding remains, 89 claw marks, 63 records from local guides, 10 records from Bantimurung Bulusaraung National Park inventory data and six calls; Fig. 1).
Model evaluation and contribution of environmental variables
The mean AUC value was 0.966 ± SD 0.006, indicating high predictive accuracy. The environmental variables tested for multicollinearity all resulted in variance inflation factor values < 10, and thus all were eligible to be used in model construction. Land-cover type had the highest contribution to the model (54.7%), followed by distance from the nearest path (22.6%), and the remaining variables each contributed < 10% (Table 1). Land-cover type was also the environmental variable with the highest contribution to the model as a single variable based on the jackknife test, with elevation, slope and NDVI also producing high AUC values (Fig. 2).
Table 1 Contribution of environmental variables to the maximum entropy model of potentially suitable habitat for the bear cuscus Ailurops ursinus in southern South Sulawesi, Indonesia (Fig. 1).

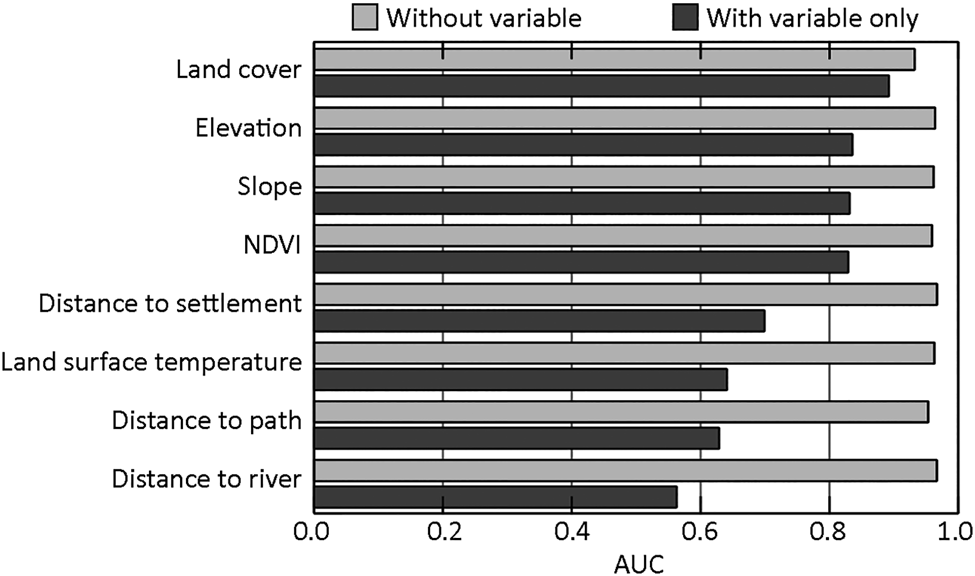
Fig. 2 Jackknife output plot to assess the predictive ability of the environmental variables for the maximum entropy (MaxEnt) model of potential bear cuscus habitat in southern South Sulawesi, Indonesia, showing the importance of individual environmental variables when used in a model relative to a model built without that variable (i.e. using the remaining environmental variables). Values are means over 10 replicate runs. AUC: area under the receiver operating characteristic curve; NDVI: normalized difference vegetation index.
Of the 18 land-cover categories used in model construction, secondary forest had the highest probability of bear cuscus presence, followed by primary forest and shrubland (Fig. 3a). The other land-cover types, which were mainly open habitats or settlement areas, had low probability of bear cuscus presence (< 0.08) and thus may either impede the species’ movement or fail to provide sufficient protection and food.
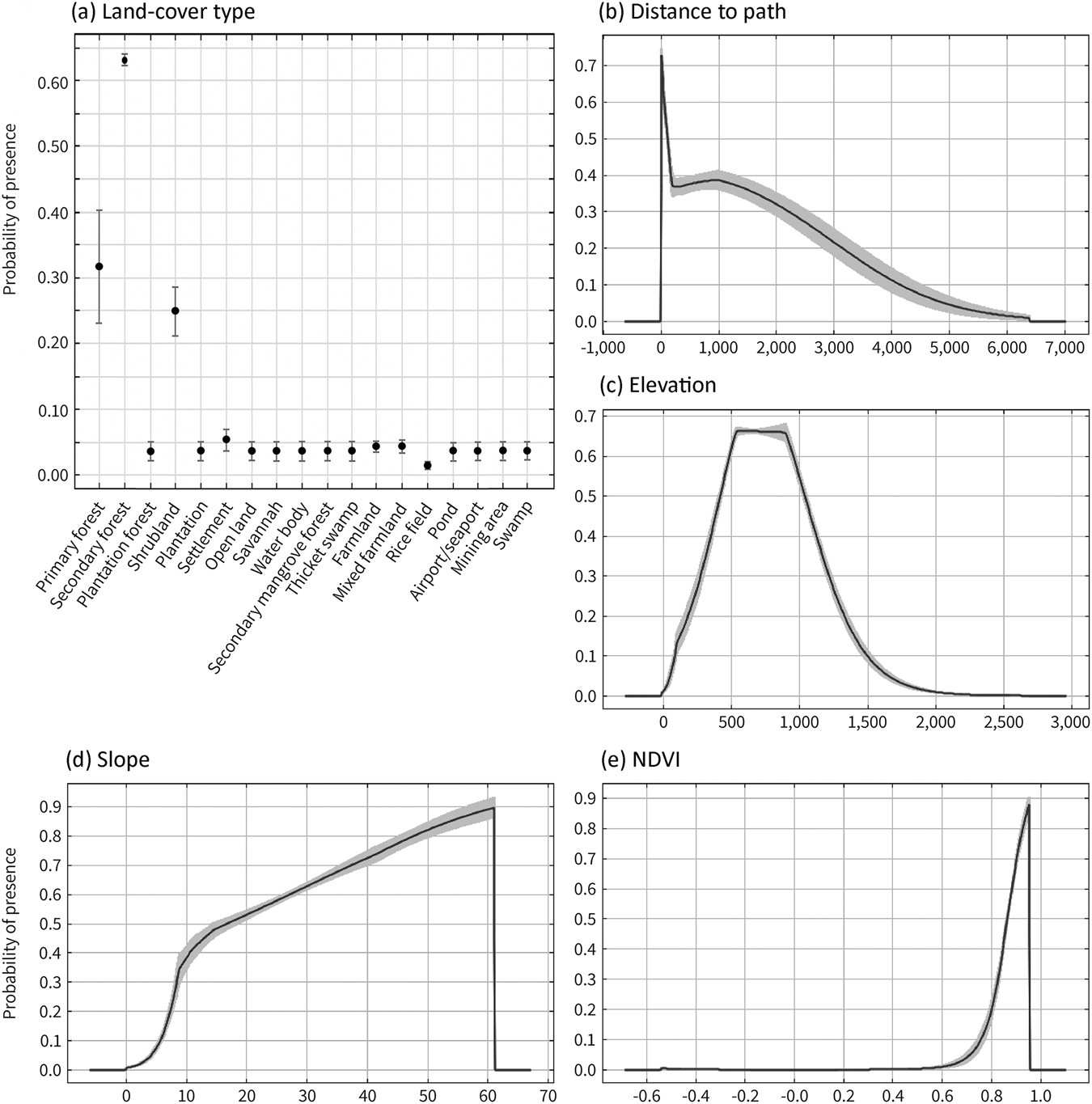
Fig. 3 Response curves of the five environmental variables with the greatest contribution to the MaxEnt model constructed for the bear cuscus habitat in southern South Sulawesi, Indonesia: (a) land-cover type, (b) distance from the nearest path, (c) elevation, (d) slope and (e) normalized difference vegetation index (NDVI). The black circles/curves show the mean response of the 10 replicate maximum entropy runs and the grey whiskers/areas the SD.
The bear cuscus was most likely to be encountered in the proximity of paths (Fig. 3b), and this probability decreased with increasing distance, although it remained relatively high up to 1,000 m from a path. The probability of presence increased with elevation until reaching the species' maximum altitude at a range of c. 500–900 m, after which the probability of presence decreased (Fig. 3c). The species was more likely to be recorded in areas with steep slopes, up to an angle of 60°, where probability of presence peaked (Fig. 3d). Areas with an NDVI value of 0.8–0.9, reflecting dense vegetation cover, were the most likely to be occupied by a bear cuscus (Fig. 3e).
Potential spatial distribution of the bear cuscus
Our modelling indicated 92.5% of the area in the southern landscape of South Sulawesi was unsuitable as bear cuscus habitat (Fig. 4, Table 2). Only 0.5 and 0.1% of the area was mostly and highly suitable, respectively, representing a total estimated area of 12,119 ha. Much of the potentially suitable habitat was found in small, often disconnected patches.
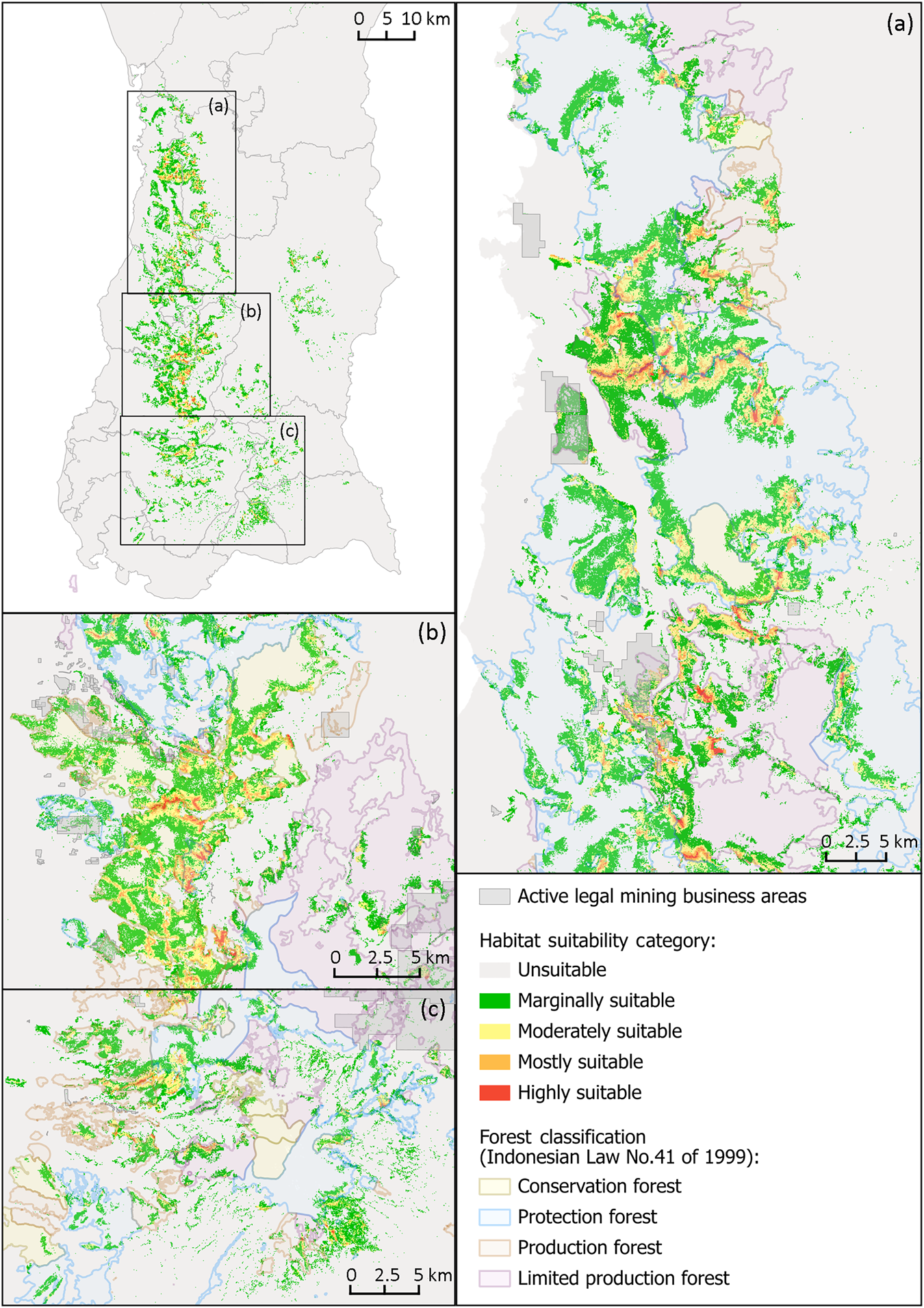
Fig. 4 Habitat suitability map of the bear cuscus in the southern landscape of South Sulawesi, Indonesia, based on our maximum entropy model, and three regions of the species’ distribution: (a) Region I, (b) Region II and (c) Region III. The map of each region shows intersections between potential bear cuscus habitats and active legal mining areas. We recommend prioritizing Regions I and II for bear cuscus conservation. (Readers of the printed journal are referred to the online article for a colour version of this figure.)
Table 2 Total area and respective percentage of bear cuscus habitat suitability in three identified regions in southern South Sulawesi, Indonesia. Region I is dominated by protection forest, Region II by conservation forest and Region III by a mixture of forest types.
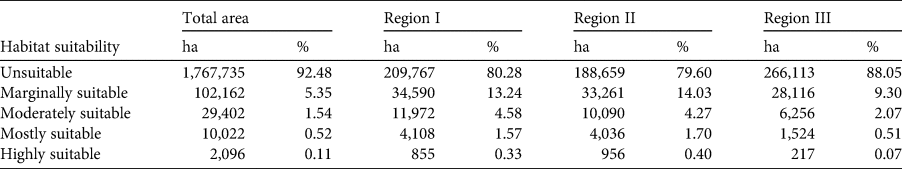
Based on the dominating designated forest function, we identified three bear cuscus distribution areas: Region I was dominated by protection forests, Region II by conservation forests, and Region III comprised a mixture of forest types. All regions had a high proportion of unsuitable habitat, ranging between 79–88% of the respective area (Table 2). In Region I (Fig. 4a), 51,525 ha was potentially suitable habitat, with a combined 4,963 ha being mostly or highly suitable, representing c. 1.9% of the region's total area. In Region II (Fig. 4b), the potentially suitable habitat totalled 48,343 ha, of which 4,992 ha was mostly or highly suitable (Table 2). Region III, consisting of forests of various functions, had the least potentially suitable habitat (Fig. 4c), totalling 36,114 ha, of which only 1,742 ha was considered mostly or highly suitable. This region also had the greatest overlap of mining areas with potentially suitable bear cuscus habitat, as well as the highest level of habitat fragmentation (Fig. 4c).
Discussion
Model evaluation
Our study had several limitations. Firstly, many parts of our research area are difficult to reach because of steep slopes, high elevation or lack of accessibility. Bantimurung Bulusaraung National Park lies across mountains up to 1,694 m tall, with difficult terrain and lack of trekking routes hampering our data collection. However, given the bear cuscus generally inhabits areas < 1,000 m, this should not affect our results. Secondly, the Park contains a part of the second largest karst ecosystem in the world, the Maros Pangkep UNESCO Global Geopark, which extends across c. 45% (19,337 ha) of its total area. The presence of karst towers, extensive cliffs and steep slopes (Duli et al., Reference Duli, Mulyadi and Rosmawati2019) within this ecosystem limited our sampling ability. Considering these limitations, and following Pearson et al. (Reference Pearson, Raxworthy, Nakamura and Peterson2007), our model should be interpreted not as an indication of the habitat requirements of the bear cuscus but as a prediction of its distribution in areas with similar environmental conditions to those in which it is known to occur.
The high predictive accuracy of the model could be attributed to the small range of the bear cuscus, as AUC values are usually high for such species (Elith et al., Reference Elith, Graham, Anderson, Dudík, Ferrier and Guisan2006; Phillips, Reference Phillips2017). The significant contribution of land-cover type to the model is a result of the species’ dependence on forest habitat (Dwiyahreni et al., Reference Dwiyahreni, Kinnaird, O'Brien, Supriatna and Andayani1999; Repi et al., Reference Repi, Masyud, Mustari and Prasetyo2020), specifically areas with sufficient cover for movement and concealment. Although primary forest is still the most important habitat for the bear cuscus (Nugraha et al., Reference Nugraha, Andriyani and Marliana2021), our results showed that the occurrence probability was highest in secondary forest, followed by primary forest and shrubland. The occurrence data also showed a comparatively high proportion of records within these three land-cover types (Supplementary Table 2). This is probably because secondary forest was the most widespread land-cover type with sufficient habitat connectivity for the bear cuscus (Fig. 5). Where a patch of secondary forest was disconnected, shrubland functioned as a corridor, connecting the forest tracts. Primary forest, however, was only found in small pockets scattered amongst secondary forest (Fig. 5). These findings are supported by Nugraha et al. (Reference Nugraha, Andriyani and Marliana2021), who found that primary forest was used by the bear cuscus only in the lowland non-dipterocarp forest inside Bantimurung Bulusaraung National Park, by only one subpopulation. Nugraha et al. (Reference Nugraha, Andriyani and Marliana2021) also highlighted that, in general, secondary forests are overused by the bear cuscus, except in lowland limestone forest, indicating that the type of secondary forest ecosystem may also contribute to the level of habitat usage. As such, we suspect the species may rely on the presence of Moraceae plants (especially figs, its preferred food source) and the intactness of canopy cover. As the bear cuscus' foraging behaviour is dictated by their need to conserve energy, they tend to forage near their resting places, and will therefore choose any habitat that provides a food source and continuous dense canopy cover.
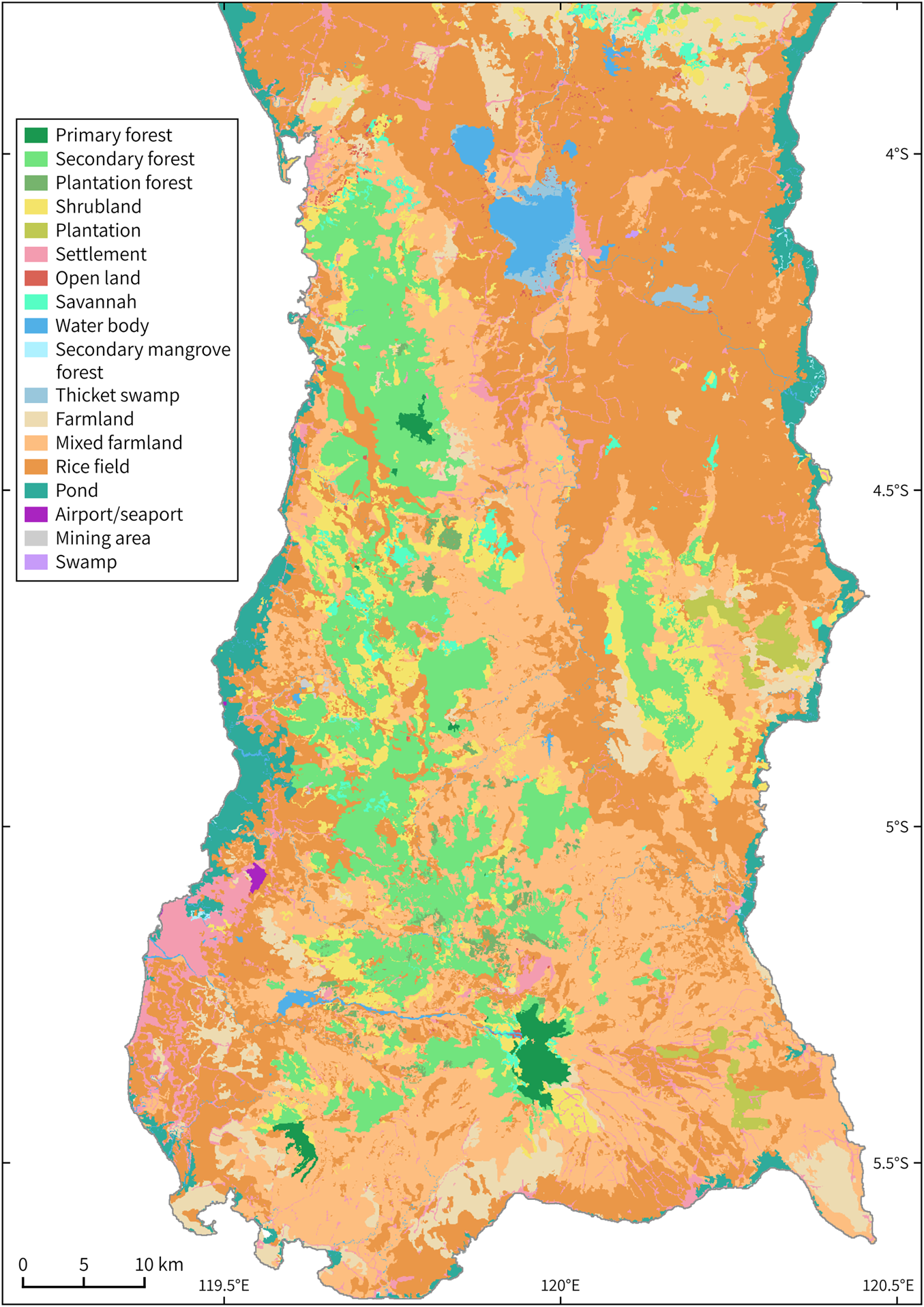
Fig. 5 Land-cover map of the modelling area in southern South Sulawesi, Indonesia. (Readers of the printed journal are referred to the online article for a colour version of this figure.)
Although the bear cuscus is sensitive to human presence (Nugraha & Mustari, Reference Nugraha and Mustari2017), our model suggests it does not necessarily avoid paths. However, the fact it was comparatively absent c. 200 m from the nearest path could be attributed to road-related disturbances, such as vehicular noise and the presence of human-made structures. The high occurrence probability near paths may therefore be a result of presence records near footpaths but not roads or highways. Overlapping tree crowns above footpaths enable the bear cuscus to move across paths whilst foraging, and so long as these paths do not disrupt its movement, the species appears not to avoid them. Consequently, this increases the chance of exposure to humans, and their susceptibility to being captured or killed. Bear cuscus encounters were more common 200–1,000 m into the forest, because of the species' preference for forest interior with interlocked canopies (Repi et al., Reference Repi, Masyud, Mustari and Prasetyo2019), where food is plentiful and human disturbance is lower.
We found the most suitable elevation for the bear cuscus was 500–900 m, aligning with previous records of the species at c. 600–1,015 m (Khairunnisa & Mustari, Reference Khairunnisa and Mustari2018), but higher than other records at 400 and 600 m (Nowak, Reference Nowak2018; Salas et al., Reference Salas, Dickman, Helgen and Flannery2019). Occurrence decreased below and above this range; at lower elevations, land-cover type was dominated by rice fields, mixed farmland and settlements (Fig. 5), and at higher elevations climatic conditions are harsher and smaller trees are more prevalent (Kartawinata, Reference Kartawinata2013) than tall trees with thick trunks preferred by the bear cuscus (Repi et al., Reference Repi, Masyud, Mustari and Prasetyo2020). Bear cuscus presence tended to increase with increasing terrain steepness, up to 60°, which was the highest slope angle within our study area. Steep slopes tend to be less accessible and more difficult to reach for both humans and predators (Nugraha et al., Reference Nugraha, Andriyani and Marliana2021), thus serving as a safe place for the species.
The probability of presence increased with increasing NDVI value, with probability highest (0.9) at the highest NDVI within the study area (0.6). As a proxy for vegetation cover (Suárez-Seoane et al., Reference Suárez-Seoane, Osborne and Rosema2004), NDVI can also be used to indicate food availability and potential habitat for arboreal marsupials (Youngentob et al., Reference Youngentob, Yoon, Stein, Lindenmayer and Held2015), and our results indicate the bear cuscus has a preference for dense vegetation cover. This concurs with previous findings showing the species tends to occupy areas with dense foliage (Repi et al., Reference Repi, Masyud, Mustari and Prasetyo2020).
Potential spatial distribution and suggested priority areas for bear cuscus conservation
Despite a large tract of forest remaining there, the southern landscape of South Sulawesi offers limited potentially suitable habitat for the bear cuscus. Given the species’ requirements for dense, interlocking canopy, it is highly probable that any anthropogenic activities that reduce habitat connectivity, even small-scale tree felling, could degrade habitat quality. Converted land around bear cuscus habitat acts as a barrier, impeding the species’ movement and limiting its ability to find food. Our results reflect how, in the absence of habitat modelling, land-use maps may create the illusion of greater habitat availability than there is, as most of the forest is not actually suitable for this arboreal marsupial. Our model showed the species' potential distribution was mainly within protection forests, which had a much greater extent than conservation forests. Although, in theory, conservation forests should provide better conditions with less disturbance, protection forests are believed to play an important role as buffer areas and natural habitats for wildlife (Khalil et al., Reference Khalil, Setiawan, Rustiati, Haryanto and Nurarifin2019), but a lack of research obscures their true value for conservation (Hero et al., Reference Hero, Hardjanto, Ahdiyar, Wijaya, Widiyanto and Wulandini2021).
Of the three regions identified as bear cuscus distribution in this study, Regions I and II hosted equally intact potential habitat and should therefore be prioritized as conservation areas for the species. However, Region II should be considered a higher priority, as it is mostly located within the conservation forest of the National Park (Fig. 4b). Similarly to other national parks, Bantimurung Bulusaraung uses a zoning management system (Pudyatmoko et al., Reference Pudyatmoko, Budiman and Kristiansen2018), with a core zone and a wilderness zone, both of which are advantageous for maintaining the integrity of bear cuscus habitat. However, in addition to these zones, our model showed there were also potentially highly suitable habitats within the utilization zone of the Park, which is publicly accessible (Broto & Setiawan, Reference Broto and Setiawan2013) and thus more exposed to anthropogenic disturbance. Further infrastructure expansion in the utilization zone is probable in the future and may compromise the availability of such habitat. Almost all of the Hasanuddin University Educational Forest area was potentially suitable bear cuscus habitat and, given its designation as an educational area, may serve as a safe haven for the species in the future. However, poaching by local people has been reported to occur frequently in and around the Park and the Educational Forest (Pado and Husain, pers. comm., 2021). Because of the threat from poaching and the substantial extent of suitable habitat in Region II, management and protection of the ecosystems in this region should be considered high priorities for conserving populations of the bear cuscus and other endemic species. In addition, although Region I had significantly greater areas of potentially suitable bear cuscus habitat in total, these were divided into two larger areas surrounded by several small patches, thus lacking habitat continuity (Fig. 4a). We recommend the forest patches in this region are connected to create corridors that facilitate bear cuscus movements. It is also important to consider that, although there is potentially suitable habitat within the conservation forest, there has been an increase in infrastructure development (primarily for tourism, e.g. food stalls, souvenir shops and public facilities), accessibility (roads and bridges; RN, pers. obs., 2021), and tourist numbers (Pagessa, Reference Pagessa2022), leading to higher intensity and frequency of disturbance experienced by the bear cuscus.
Overall, Region III offered the least valuable habitat for the bear cuscus and should be the lowest priority for conservation. Here, patches of suitable habitat were found in secondary forest, separated by 2–30 km of unsuitable habitat, a distance too great for the bear cuscus to traverse safely. Many parts of this region are also > 1,000 m, an altitude at which the species is rarely recorded (Nugraha et al., Reference Nugraha, Andriyani and Marliana2021). The bear cuscus population is therefore isolated into smaller local subpopulations, impeding gene flow. Our model predicted a small extent of suitable habitat within the region's four conservation forests. Two of these are located on the mountain slopes and mostly composed of montane pine forest at 1,000–2,300 m, making bear cuscus presence unlikely because of the high elevation and the limited value of pine trees for food or shelter. This region also had the greatest overlap of suitable habitat with mining areas (gold and iron ore), further limiting the potentially suitable habitat. In this region we recommend increasing canopy cover and connectivity between habitat patches, and minimizing mining activities within potential bear cuscus habitat.
Management of potential bear cuscus habitat: minimizing threats and maximizing protection
Our findings indicate the bear cuscus should be considered for legal protection because of habitat fragmentation caused by settlement expansion and resource exploitation. In addition to its northwards dispersal being cut off, the high proportion of potentially unsuitable habitat in the modelling area serves as a warning for current management practices in South Sulawesi. Our model predicted that areas with frequent poaching in the National Park and the Educational Forest in Region II qualify as potentially suitable bear cuscus habitat, and these areas are also part of the species’ current range. In many parts of our study area the bear cuscus has been illegally hunted for consumption, trade between villages (e.g. for chickens), rearing or pet food (Tingkang and I. Ardi, pers. comm., 2021). Previous reports also indicate widespread knowledge of hunting for consumption and trade in Hasanuddin University Educational Forest (Putri et al., Reference Putri, Maulany and Achmad2022). Furthermore, there continues to be consistent demand for bear cuscus meat at bushmeat markets (Bailey et al., Reference Bailey, Hilser, Siwi, Lawe, Waterman and Loffeld2022). Stronger action must be taken to combat poaching, including developing alternative economic opportunities for local people, improving awareness and environmental education, and mitigating demand for bushmeat.
Habitat loss from land conversion is a significant threat to the bear cuscus, particularly as licensed mining operations continue to increase (Saing et al., Reference Saing, Djainal and Deni2021). In Region II, many of the mining operations are within potential bear cuscus habitat (Fig. 4b). These operations, which mainly focus on limestone and marble, often employ harmful extractive processes (e.g. the use of explosives), that cause substantial damage to the surrounding ecosystem (Duli et al., Reference Duli, Mulyadi and Rosmawati2019; Fatinaware et al., Reference Fatinaware, Fauzi and Hadi2019). As well as the expansive activities of a cement company (Duli et al., Reference Duli, Mulyadi and Rosmawati2019), land used for mining increased six-fold during 2005–2019 (Saing et al., Reference Saing, Djainal and Deni2021), and mining concessions in South Sulawesi will probably continue to expand. Outside the legal concessions, small-scale mining by local people is reportedly widespread (Fatinaware et al., Reference Fatinaware, Fauzi and Hadi2019). Not only could the proximity to potentially suitable bear cuscus habitat be associated with further degradation, but the prevalence of mining suggests our results are conservative, as illegal mining activities have been found to cause extensive deforestation in the Amazon (Siqueira-Gay & Sánchez, Reference Siqueira-Gay and Sánchez2021), pollute water and reduce soil fertility in Ghana (Obeng et al., Reference Obeng, Oduro, Obiri, Abukari, Guuroh and Djagbletey2019) and affect gorilla and chimpanzee populations in the Democratic Republic of the Congo (Spira et al., Reference Spira, Kirkby, Kujirakwinja and Plumptre2019). In Sulawesi, more work is needed to examine the ecological impacts of these illegal activities.
In addition to legal protection by the Indonesian Government, we further recommend that the bear cuscus is considered for reassessment on the IUCN Red List, where it is categorized as Vulnerable (Salas et al., Reference Salas, Dickman, Helgen and Flannery2019). Since the latest assessment in 2015, its legal status has changed (it is no longer protected), as have the areas in which it occurs. In North Sulawesi, part of a project said to be the largest infrastructure programme in human history is underway (Ng et al., Reference Ng, Campos-Arceiz, Sloan, Hughes, Tiang, Li and Lechner2020), reducing what is already the smallest forest area in Sulawesi; and in Central Sulawesi it is predicted 20% of forest cover will be lost within the next 10 years (Voigt et al., Reference Voigt, Supriatna, Deere, Kastanya, Mitchell and Rosa2021). In the focal area of our study, 125,707 ha (12.5%) of forest cover was lost during 2000–2023, 72,952 ha of which since 2015 (Global Forest Change data; Hansen et al., Reference Hansen, Potapov, Moore, Hancher, Turubanova and Tyukavina2013), indicating that deforestation has been increasing rapidly in recent years (Supplementary Fig. 2). Our findings suggest only a portion of the protected areas reported previously (Salas et al., Reference Salas, Dickman, Helgen and Flannery2019) can provide suitable refuge for the bear cuscus, as they are predominantly mountainous and face the same illegal activities noted in our research area (Supriatna et al., Reference Supriatna, Shekelle, Fuad, Winarni, Dwiyahreni and Farid2020). Hunting of the species occurs widely, and there appears to be a lack of concern for its welfare (Putri et al., Reference Putri, Maulany and Achmad2022). Mining represents a significant current and future threat as more concessions open within suitable bear cuscus habitat. Our study highlights the limited representation of South Sulawesi's bear cuscus populations in existing studies, which is of additional concern now they lack dispersal pathways beyond South Sulawesi, further isolating them from populations in the northern part of the island, where research has predominantly been focused. This is probably also true of other forest-dwelling species in the region. Further investigation may shed light on whether the bear cuscus can serve as a Cinderella species (Smith et al., Reference Smith, Veríssimo, Isaac and Jones2012), as its habitat requirements mean its conservation may benefit other species. More research is needed on its connections to the sympatric Sulawesi dwarf cuscus, Makassar tarsier and moor macaque. In Bantimurung Bulusaraung National Park and Hasanuddin University Educational Forest, we found the Makassar tarsier to prefer similar tree species to the bear cuscus, albeit in different growth forms (Andriyani et al., Reference Andriyani, Nugraha and Marliana2021).
Increasing landscape connectivity is imperative. In Hasanuddin University Educational Forest, restoration that incorporates fast-growing tree species has been suggested to meet the needs of both local fauna and people (Riley et al., Reference Riley, Albani, Zak, Germani, Rothman, Carosi and Ngakan2023). However, these trees are unsuitable for canopy-dwelling species, as our findings showed; high canopies are an important habitat requirement for the bear cuscus, which forages and rests mostly in large rainforest trees (e.g. Alstonia scholaris, Pterocarpus indicus, Ficus sumatrana), some of which are native or endemic to Sulawesi, such as Myristica impressa and Terminalia supitiana (Nugraha et al., Reference Nugraha, Andriyani and Marliana2021). Our findings broadly imply that restoration may be insufficient to protect shy, upper-canopy-dwelling species (especially those with specific behaviours, such as movement requiring continuous canopy cover), and prevention of habitat degradation is a more effective measure (i.e. maintaining rather than replacing resources). Although the bear cuscus can adapt to secondary forest and does not depend solely on primary forest, this may expose it to increased anthropogenic threats and less nutritious food sources, compromising its well-being and reproduction. As such, the first steps in a conservation plan must be to slow the expansion of resource extraction and human settlement and protect what remains of the fragmented landscape to limit the decline of one of Sulawesi's lesser-known endemic species.
Author contributions
Study design: both authors; fieldwork: RN; data analysis, writing: both authors.
Acknowledgements
We thank the management of Bantimurung Bulusaraung National Park and the Faculty of Forestry, Hasanuddin University, for providing their permission and facilities during the research; the local guides of Bantimurung Bulusaraung National Park for their assistance in the field, namely Mr Pado at Pattunuang Resort, Irvan Ardi at Balocci Resort, Mr Tingkang at Tondong Tallasa Resort and Mr Haji Ismail, Mr Parman and Mr Azis at Mallawa Resort, as well as Mr Husain at Hasanuddin University Educational Forest; our colleagues from Alauddin State Islamic University of Makassar and Makassar Forestry Vocational High School for assisting with the data collection; and Joaquim Baeta for editing the manuscript.
Conflicts of interest
None.
Ethical standards
This research abided by the guidelines on ethical standards of both Oryx and the government of Indonesia, and appropriate field protocols and research permissions were obtained. No bear cuscus individuals were handled and no specimens were collected, and surveys were conducted with the permission of the respective landowners.
Data availability
The data that support the findings of this study are partially available on Zenodo at doi.org/10.5281/zenodo.12736845, doi.org/10.5281/zenodo.12736841 and doi.org/10.5281/zenodo.12737698. The full data are not openly available because of the species’ vulnerability to poaching, but are available from the corresponding author upon reasonable request.










 Open access
Open access