



1. Introduction
The Ediacaran Period (635–538.8 Ma) records the rise of complex macroscopic life and the oldest recognized animals (metazoans) in the fossil record. The first fossils that are widely considered to represent metazoan taxa are found at <580 Ma, with putative metazoan trace fossils appearing by at least 560 Ma, and the first skeletal taxa at ca. 550 Ma (e.g. Germs, Reference Germs1972; Jensen, Reference Jensen2003; Matthews et al. Reference Matthews, Liu, Yang, McIlroy, Levell and Condon2020; Bowyer et al. Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b).
The phylogenetic affinities of the Ediacara biota are unclear, but they probably include stem- and early crown-group animals, as well as possible non-metazoans (Erwin et al. Reference Erwin, Laflamme, Tweedt, Sperling, Pisani and Peterson2011; Dunn et al. Reference Dunn, Liu, Grazhdankin, Vixseboxse, Flannery-Sutherland, Green, Harris, Wilby and Donoghue2021). The Ediacara biota has been informally divided into the Avalon, White Sea and Nama assemblages, but these assemblages are often (but not always) based on soft-bodied macrofossils alone so excluding skeletal taxa, trace fossils, or embryo-like microfossils. Available radiometric ages and global compilations of biotic distributions for this interval, in addition to network analyses, confirm that these represent temporally overlapping but broadly successive assemblages (Grazhdankin, Reference Grazhdankin2004; Boag et al. Reference Boag, Darroch and Laflamme2016; Muscente et al. Reference Muscente, Bykova, Boag, Buatois, Mángano, Eleish, Prabhu, Pan, Meyer, Schiffbauer, Fox, Hazen and Knoll2019; Yang et al. Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonaldn and Zhu2021). The distribution of assemblages may also have been controlled by differences in sedimentary setting and relative water depth, palaeoecology or taphonomy (e.g. Grazhdankin, Reference Grazhdankin2004; Narbonne, Reference Narbonne2005; Boag et al. Reference Boag, Darroch and Laflamme2016). It has been proposed, therefore, that these assemblages might better be termed palaeocommunities (i.e. recurrent and distinguishable associations of taxa), which inhabited different environmental settings but where extinctions nonetheless shaped their evolutionary dynamics (Muscente et al. Reference Muscente, Bykova, Boag, Buatois, Mángano, Eleish, Prabhu, Pan, Meyer, Schiffbauer, Fox, Hazen and Knoll2019).
An ecological shift in feeding mode and life habit accompanies the increase in generic diversity from the Avalon to White Sea assemblages (Evans et al. Reference Evans, Tu, Rizzo, Surprenant, Boan, McCandless, Marshall, Xiao and Droser2022). Both the Avalon and White Sea assemblages largely disappeared by ca. 550 Ma (Grazhdankin, Reference Grazhdankin2014; Yang et al. Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonaldn and Zhu2021), where analysis suggests that taxa with a high surface area to volume ratio may have preferentially survived, inferred to potentially reflect increased resilience to low environmental oxygen levels (Evans et al. Reference Evans, Tu, Rizzo, Surprenant, Boan, McCandless, Marshall, Xiao and Droser2022). Geochemical proxies, however, suggest that this extinction pre-dates a decrease in global oxygen availability (Tostevin et al. Reference Tostevin, Clarkson, Gangl, Shields, Wood, Bowyer, Penny and Stirling2019). A further decline in biodiversity is proposed to have occurred at ca. 545 Ma, although the precise timing of this decline remains uncertain (Grazhdankin, Reference Grazhdankin2014; Boag et al. Reference Boag, Darroch and Laflamme2016; Muscente et al. Reference Muscente, Bykova, Boag, Buatois, Mángano, Eleish, Prabhu, Pan, Meyer, Schiffbauer, Fox, Hazen and Knoll2019). A distinctive global, cosmopolitan, assemblage consisting of a lower diversity of existing soft-bodied morphogroups (largely erniettomorphs with subordinate rangeomorphs, arboreomorphs and dickinsoniomorphs) persisted, with new diverse skeletal, and non-skeletal tubular fossils, and more complex traces appearing <550 Ma and continuing until at least 539 Ma (Smith et al. Reference Smith, Nelson, Tweedt, Zeng and Workman2017; Nelson et al. Reference Nelson, Ramezani, Almond, Darroch, Taylor, Brenner, Furey, Turner and Smith2022). Network analysis has confirmed this to be a statistically significant grouping separate from all other Ediacaran taxa, hence named the Terminal Ediacaran biozone, and further proposed to correspond to the Nama palaeocommunity (Muscente et al. Reference Muscente, Bykova, Boag, Buatois, Mángano, Eleish, Prabhu, Pan, Meyer, Schiffbauer, Fox, Hazen and Knoll2019). The Nama Assemblage was originally named after the biota found in the Nama Group, Namibia (≥550.5 to <538 Ma), where soft-bodied fossils are reported to extend from the Kliphoek Member of the Dabis Formation through the entire overlying succession until the lower Nomtsas Formation (Waggoner, Reference Waggoner2003; Nelson et al. Reference Nelson, Ramezani, Almond, Darroch, Taylor, Brenner, Furey, Turner and Smith2022).
The Nama Group preserves a rich biota which typifies the late Ediacaran Nama Assemblage and offers one of the best palaeontological records of the terminal Ediacaran. The first soft-bodied macroscopic fossils from the Nama Group were described in the early 20th century (e.g. Gürich Reference Gürich1930, Reference Gürich1933), and since then, a diverse array of soft-bodied and calcifying macrofossils, trace fossils and microbial structures has been reported (e.g. Germs, Reference Germs1972; Narbonne et al. Reference Narbonne, Saylor and Grotzinger1997; Grotzinger et al. Reference Grotzinger, Watters and Knoll2000; Jensen et al. Reference Jensen, Saylor, Gehling and Germs2000; Noffke, et al. Reference Noffke, Knoll and Grotzinger2002; Darroch et al. Reference Darroch, Cribb, Buatois, Germs, Kenchington, Smith, Mocke, O’Neil, Schiffbauer, Maloney, Racicot, Turk, Gibson, Almond, Koester, Boag, Tweedt and Laflamme2021). Calcified taxa are largely limited to carbonate strata, with Cloudina known from ca. 550 to 551 Ma (Bowring et al. Reference Bowring, Grotzinger, Condon, Ramezani, Newall and Allen2007; Bowyer et al. Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b) and persisting until 538.56 +0.08/−0.09 Ma (Nelson et al. Reference Nelson, Ramezani, Almond, Darroch, Taylor, Brenner, Furey, Turner and Smith2022). Soft-bodied taxa belong mainly to either the Rangeomorpha or Erniettomorpha, known from the siliciclastic Kliphoek Member of the upper Dabis Formation in the Kuibis Subgroup, and Spitskop Member within the Schwarzrand Subgroup, and are widely distributed throughout the Witputs Sub-basin. Aspidella holdfasts have also been reported from the siliciclastic Nudaus Formation in the Zaris Sub-basin (Darroch et al. Reference Darroch, Boag, Racicot, Tweedt, Mason, Erwin and Laflamme2016). Both the oldest trace and body fossils known from the Nama Group have been reported from the Kliphoek Member (Darroch et al. Reference Darroch, Cribb, Buatois, Germs, Kenchington, Smith, Mocke, O’Neil, Schiffbauer, Maloney, Racicot, Turk, Gibson, Almond, Koester, Boag, Tweedt and Laflamme2021; Fig. 1c).
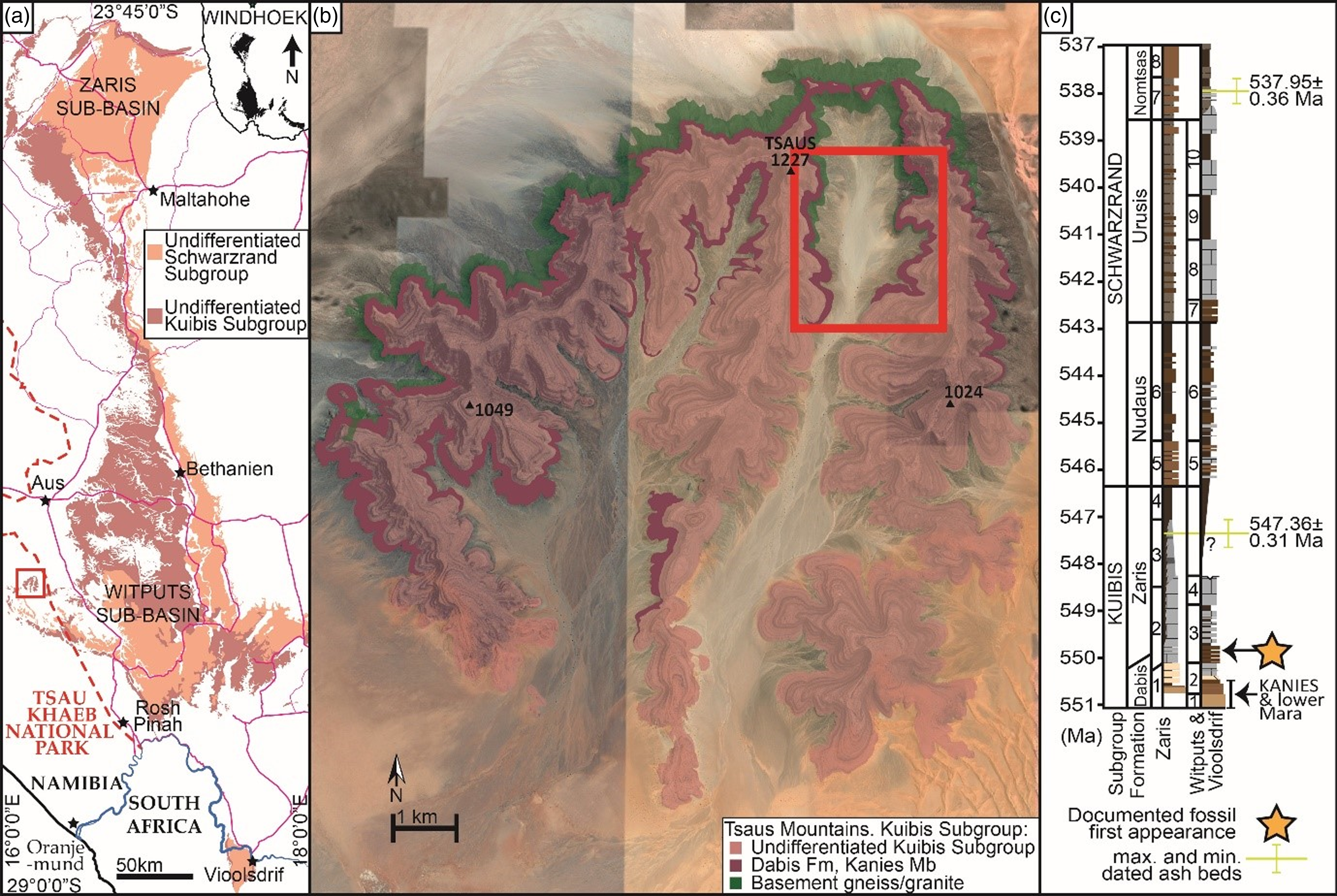
Figure 1. (a). Map of Nama Group, Namibia. (b). Geological map of the Tsaus Mountains superimposed upon Google Earth image, with box showing the outcrop of Kanies and lower Mara Members studied. (c). General Nama stratigraphy, with minimum and maximum age of dated beds and previously documented first appearance of fossil biota. Zaris Sub-basin: 1, Kanies Member; 2, Omkyk Member; 3, Hoogland Member; 4, Urikos Member; 5, Niederhagen Member; 6, Vingerbreek Member; 7, Kreyrivier Member; 8, Niep Member. Witputs and Vioolsdrift sub-basins: 1, Kanies Member; 2, Mara Member; 3, Kliphoek Member; 4, Mooifontein Member; 5, Niederhagen Member; 6, Vingerbreek Member; 7, Nasep Member; 8, Huns Member; 9, Feldschuhhorn Member; 10. Spitskop Member. Star marks previous oldest documented fossils.
Here we present a preliminary description of a potential biota from the lower Mara Member of the lower Dabis Formation from a new site, the Tsaus Mountains, thus extending the oldest known macrofossil record of the Nama Group. The Tsaus Mountains outcrop in the Tsau Khaeb National Park (formerly the Sperrgebiet, or diamond-mining ‘Prohibited Area’) and represent the western-most outcrop of the Nama Group in the Witputs Sub-basin investigated to date (Fig. 1b). We compare this biota with broadly coeval global sections and using a new global age model consider the distribution of Ediacaran taxa to offer further insight into the evolution of Ediacaran assemblages.
2. Geological setting
The Nama Group is a mixed carbonate and siliciclastic foreland basin succession deposited in supratidal to outer ramp settings in two sub-basins, the northern ‘Zaris’ and southern ‘Witputs’ (Fig. 1a; Germs, Reference Germs1983; Germs and Gresse, Reference Germs and Gresse1991). The Zaris and Witputs sub-basins deepened to the north/northwest and southwest, respectively, during deposition of the Kuibis Subgroup, with increasing distance from an intervening palaeobathymetric high (the Osis Arch), and from the Kalahari Craton to the present east (Fig. 1a). Strata within both sub-basins have been correlated using litho-, chemo- and chronostratigraphy (Fig. 1c) (Germs, Reference Germs1983; Saylor et al. Reference Saylor, Grotzinger and Germs1995, Reference Saylor, Kaufman, Grotzinger and Urban1998; Wood et al. Reference Wood, Poulton, Prave, Hoffmann, Clarkson, Guilbaud, Lyne, Tostevin, Bowyer, Penny, Curtis and Kasemann2015; Nelson et al. Reference Nelson, Ramezani, Almond, Darroch, Taylor, Brenner, Furey, Turner and Smith2022). The Nama Group consists of the dominantly Ediacaran Kuibis and Schwarzrand subgroups, and Cambrian Fish River Subgroup. Siliciclastic sediments were largely sourced from the Kalahari Craton during deposition of the Kuibis Subgroup, but were supplemented by the Damara orogenic belt to the north during deposition of the Schwarzrand Subgroup (Germs, Reference Germs1972, Reference Germs1983).
The age of the base of the Nama Group remains uncertain, but is estimated to be ≥550.5 Ma based on inferred rates of deposition for strata that underlie a dated ash bed (547.36 ± 0.31 Ma) in the lower Hoogland Member (Kuibis Subgroup) of the Zaris Sub-basin (Saylor et al. Reference Saylor, Kaufman, Grotzinger and Urban1998; Bowring et al. Reference Bowring, Grotzinger, Condon, Ramezani, Newall and Allen2007; Bowyer et al. Reference Bowyer, Zhuravlev, Wood, Shields, Zhou, Curtis, Poulton, Condon, Yang and Zhu2022).
The Kuibis Subgroup is divided into the Dabis and overlying Zaris formations (Germs, Reference Germs1983; Saylor et al. Reference Saylor, Grotzinger and Germs1995). In the Witputs Sub-basin, the Dabis Formation (Kanies, Mara, Kliphoek and Aar members) consists of two incomplete third-order sequences. The first sequence is represented by the Kanies and Mara members, and the Kliphoek and Aar members with the overlying Mooifontein Member of the Zaris Formation form the second third-order sequence. Coarse siliciclastics of the Kanies and lower Kliphoek members mark the lowstand deposits, and the Mara, Aar and Mooifontein members consist of fine siliciclastics and carbonates forming transgressive to highstand deposits (Fig. 1c; Saylor et al. Reference Saylor, Grotzinger and Germs1995; Hall et al. Reference Hall, Kaufman, Vickers-Rich, Ivantsov, Trusler, Linnemann, Hofmann, Elliott, Cui, Fedonkin, Hoffmann, Wilson, Schneider and Smith2013). Sequence boundaries are commonly overlain by coarse siliciclastic lowstand deposits in more proximal, eastern exposures, but correlative conformities complicate the lateral association of member boundaries in deeper, more continuous sections to the west (Saylor et al. Reference Saylor, Grotzinger and Germs1995).
The Tsaus Mountains cover an area of ca. 11 km2 within the Tsau Khaeb National Park. The succession is 10 km to the northwest of the described section on Farm Grens (Wood et al. Reference Wood, Poulton, Prave, Hoffmann, Clarkson, Guilbaud, Lyne, Tostevin, Bowyer, Penny, Curtis and Kasemann2015) and 50 km to the southwest of the Farm Aar section (Fig. 1b). In the Witputs Sub-basin, the lower Kuibis Subgroup was deposited diachronously from west to east during initial transgression onto the Proterozoic basement (Germs, Reference Germs1983; Saylor et al. Reference Saylor, Grotzinger and Germs1995). The Tsaus Mountains succession therefore preserves some of the oldest Nama Group strata of the Dabis Formation (Bowyer et al. Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b).
In most places, the Kanies Member (Mb) nonconformably overlies a variably metamorphosed Proterozoic granitic basement and records lowstand deposition in dominantly fluvial, and occasional marginal-marine, depositional settings (Germs, Reference Germs1983; Saylor et al. Reference Saylor, Grotzinger and Germs1995). The base of the overlying Mara Mb is marked by the initiation of a marine transgression that progressed from west to east across the underlying Kanies Mb clastics in distal settings or the planar basement contact in more proximal settings (Saylor et al. Reference Saylor, Grotzinger and Germs1995). Kanies and lower Mara Mb deposits fine upwards from very coarse, sometimes pebbly, massive sandstones (Kanies Mb) to bedded sandstones, siltstones and minor carbonate interbeds (lower Mara Mb), before transitioning to carbonate-dominated strata (upper Mara Mb) that record continued transgression across the Witputs Sub-basin (Saylor et al. Reference Saylor, Grotzinger and Germs1995). The Kanies Mb thickens to the west and south, consistent with increasing accommodation space, and reaches a maximum thickness of ∼95 m at Farm Witputs, >70 km to the south of the Tsaus Mountains (Saylor et al. Reference Saylor, Grotzinger and Germs1995). The lower Mara Member at both Tsaus and Grens were thus deposited during a diachronous transgression, and this interval is associated with subaerial exposure and non-deposition or non-preservation due to erosion at more proximal, easterly sections. No macrofossil biota have been recorded from siliciclastics of the Kanies Mb or lower Mara Mb to date at any Nama Group localities.
3. Results
In the area studied within the Tsaus Mountains (Fig. 1b), the Kanies Mb and lower Mara Mb together reach ca. 61 m in thickness (Fig. 2a, b). The lowermost beds (0 – 1.5 m) deposited upon the basement contact consist of planar to cross-bedded very coarse, friable, yellow sandstones (Fig. 2c). The succeeding ca. 25.5 m interval is often cliff-forming and consists of 0.5 – 1.5 m cycles of massive to planar-bedded decimetre scale beds of coarse- to medium-grained sandstones.

Figure 2. (a), Sedimentary log of Kanies and lower Mara Members, with enlargement of Fig. 1b, showing location of images. (b, f) Kanies Member and transgressive surface that marks the base of the overlying Mara Member (dashed line). (c) Contact of base of Kanies Member with Proterozoic basement granite. (d) Asymmetric ripples on bedding surface in the lower Mara Mb. (e) Interference ripples in the lower Mara Mb. (f) Lower Mara Member showing cyclicity, and position of first appearance of dolostone relative to the basal Mara transgressive surface (dashed line).
Overlying this is a ca. 16 m-thick interval consisting of four coarsening-up cycles of medium to fine sandstone. We provisionally assign the boundary between the Kanies and Mara members to the base of this interval at 27 m based on the defining characteristics of the original Member subdivision (Saylor et al. Reference Saylor, Grotzinger and Germs1995; Bowyer et al. Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b). In these cycles, most beds have sharp planar bases and bedding plane surfaces that commonly exhibit straight-crested, sinuous or bifurcating symmetrical ripples (Fig. 2d), and less common asymmetrical and interference ripples (Figs. 2e, 3f). The succession continues to fine upwards in the interval 43 – 61 m, with nine further cycles of fine sandstone and siltstone (Fig. 2f). Here the sandstone units, with undulating bases, become progressively thinner and the siltstone units thicker, with the thickest siltstone unit occurring at ca. 55 m. Two thin (<0.2 m) buff-coloured dolostone beds, the first discontinuous and the second laterally continuous, appear at ca. 56 m and 57 m, respectively (Fig. 2a). The dominant lithology of the Mara Mb switches at ca. 61 m to medium-bedded mid- to dark-grey dolostone (Fig. 2a, b, f).

Figure 3. Potential body and trace fossils of the lower Mara Mb (a), in situ rippled bed surface with possible Bergaueria or holdfasts, one with a juvenile rangeomorph or arboreomorph frond, and Horodyskia. (b) Possible Horodyskia, or trace fossil on bed surface. (c) Possible juvenile rangeomorph or arboreomorph frond and associated holdfast on bed surface. (d) Possible Beltanelliformis or Intrites on float, but unclear as to whether bed top or bed sole surface. (e), enlargement of (d). (f) Possible Beltanelliformis or Intrites on asymmetrical ripples on bed top. (g)–(k) Float samples and unclear as to whether bed top or bed sole surface. (g) Possible Beltanelliformis. (h) Possible Beltanelliformis or Intrites. (i) Possible Beltanelliformis or Intrites. (j) Possible Beltanelliformis, with separate, multiple depressions within each domal structure (see insets). (k) Elongate and sinuous, possibly ridged, structures. Scale bars = 10 mm.
3.a. Biota
At ca. 44 m above the basement contact, an in situ, symmetrically rippled medium-grained sandstone bed top surface is exposed which preserves numerous structures that may be biotic in origin (Fig. 3a). These include a number of circular forms, 5 – 10 mm in diameter (Fig. 3a, b). These are preserved as positive epirelief above the bedding surface and delineated by a sharp edge.
While some forms are isolated, around nine individual structures of diameter ca. 5 mm are arranged in a sinusoidal pattern on the bedding surface to form a variably continuous chain (Fig. 3a, b). This chain appears to change orientation depending on the part of the ripple surface occupied. Two segments of the chain are preserved between the crests of ripples and are aligned parallel to each other, and the ripple trough. When the structure is preserved on the crest of a ripple, the chain orientation changes by ∼90 degrees, that is perpendicular to the ripple crest (Fig. 3b).
One of the raised circular features exhibits apparent continuity with an elongated element that displays a potentially frondose morphology (Fig. 3a, c). This elongate element is ca. 40 mm in length and characterized by a straight axis that extends from the centre of the circular feature. A minimum of five closely spaced, raised teardrop-shaped elements, arranged en-echelon, extend ca. 8 mm from one side of this axis (Fig. 3c). Near the terminal end of the axis, furthest from the circular feature, a series of teardrop-shaped elements radiate out from the axis, with the longest terminal element aligned near-parallel to the axis (Fig. 3c). Both the raised circular feature and elongate frondose element are distinguished from the surrounding sediment by a thin dark brown to black mineral veneer (Fig. 3a, c).
A number of sandstone float samples preserve surfaces that are populated with further raised, circular to oblong features (Fig. 3d–f). These are frequently domed and vary in size, with the largest being ca. 25 mm in diameter and the smallest being ca. 5 mm in diameter. These features are found in greater abundance than those described above from the specimen at ca. 44 m, but individual features are separated from one another despite displaying a higher density on the bedding surface. Whether these structures are found on a bed top or bed sole surface is not clear (Fig. 3d, e), but in other cases, coarser grains infilling ripple troughs show that these are preserved in positive epirelief on rippled bed top surfaces (e.g. Fig. 3f).
Further sandstone float samples show broadly circular and closely spaced, raised structures (Fig. 3g–j). It is unclear as to whether these are found on bed top or bed sole surfaces. Some have diameters of between ca. 10 and 25 mm, are raised above the surface of the sample and have a sharp edge, with interior surfaces that are flattened to dome-shaped (Fig. 3g). Other specimens that are characterized by a circular to oblong morphology commonly have maximum diameters of ca. 5 – 10 mm (Fig. 3h, i). Many of the structures have poorly defined outlines (e.g. Fig. 3e, h). One specimen populated by flattened circular features with well-defined sharp edges hosts some individuals with central depressions that appear, in some cases, to exhibit some consistency in shape, and are more complex than simply being circular or oval (Fig. 3j, insets).
Some float samples preserve a sinuous, raised form (again unclear as to whether on bed top or sole), that is populated with raised ridges, transversely oblique to the long axis of the structure (Fig. 3k). The structure is ca. 50 mm in length and of variable width, from 3 to 8 mm, where each ridge is ca. 1 mm wide. The form appears to taper to a point at one end.
Other float samples, and similar samples found in situ on bed tops at ca. 40 – 42 m above the basement contact, display sinuous forms on their upper surfaces, preserved in positive epirelief, and potentially counterparts on lower surfaces of separate samples (Fig. 4). Some of the structures preserve first- and second-order sinusoidal forms, meandering across the surface of the samples (Fig. 4a, c, e). The raised structures are ca. 1 cm in diameter, have a curved upper surface and are laterally discontinuous. An individual form may extend up to ca. 10 – 15 cm (Fig. 4a) or as little as ca. 4 – 5 cm (Fig. 4b), abutting at a curved edge. The structures are separated from each other; however, some may be tenuously connected by thin, raised regions (Fig. 4i).

Figure 4. Sedimentary surface textures, possibly wrinkle structures known as ‘Kinneyia’ texture, on bed top surfaces of the lower Mara Mb. Scale bars = 10 cm.
Many in situ and float specimens preserve raised, lozenge-shaped features on bed tops that can either be isolated (Fig. 5e) or form short chains with offsets between each ‘lozenge’ (Fig. 5a, b). ‘Lozenges’ can be ca. 1 – 5 mm wide and reach a length of ca. 10 mm. Longer chains composed of individual lozenges can reach lengths of up to ca. 10 cm and are often aligned parallel to other chains, almost completely covering the bedding surface (e.g. Fig. 5c, f).

Figure 5. Pseudofossils of the lower Mara Mb on bed top surfaces, formed by infill of shrinkage cracks or en-echelon lozenge-shaped crack structures. (a), (b), (d) Small-chain-like features of one to four structures; (e) isolated structures; (c), (f) continuous, aligned, chains. Scale bars = 10 mm.
4. Discussion
4.a. Sedimentological setting and Stratigraphy
In the vicinity of Aus, the Kanies and lower Mara members combined are thickest in the most distal sections, in the Tsaus Mountains and Grens 1, where accommodation space was greatest (Fig. 6; Wood et al. Reference Wood, Poulton, Prave, Hoffmann, Clarkson, Guilbaud, Lyne, Tostevin, Bowyer, Penny, Curtis and Kasemann2015). In the section reported to the south at Farm Witputs, the Kanies Mb is thicker than at Tsaus (Saylor et al. Reference Saylor, Grotzinger and Germs1995). High-frequency cycles are also preserved above the transgressive surface that marks the boundary between the Kanies and Mara members, in both the Tsaus Mountains and at Grens 1, where coarse and medium-grained sandstone, and medium-grained sandstone and siltstone interbeds provide evidence for short-term sea level cycles (Fig. 6). The remainder of the Mara Mb is composed of evaporative-dolostone units followed by limestones (grainstones) in all sections, but notably condensed to the east at Arasab (Fig. 6; Bowyer et al. Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b).
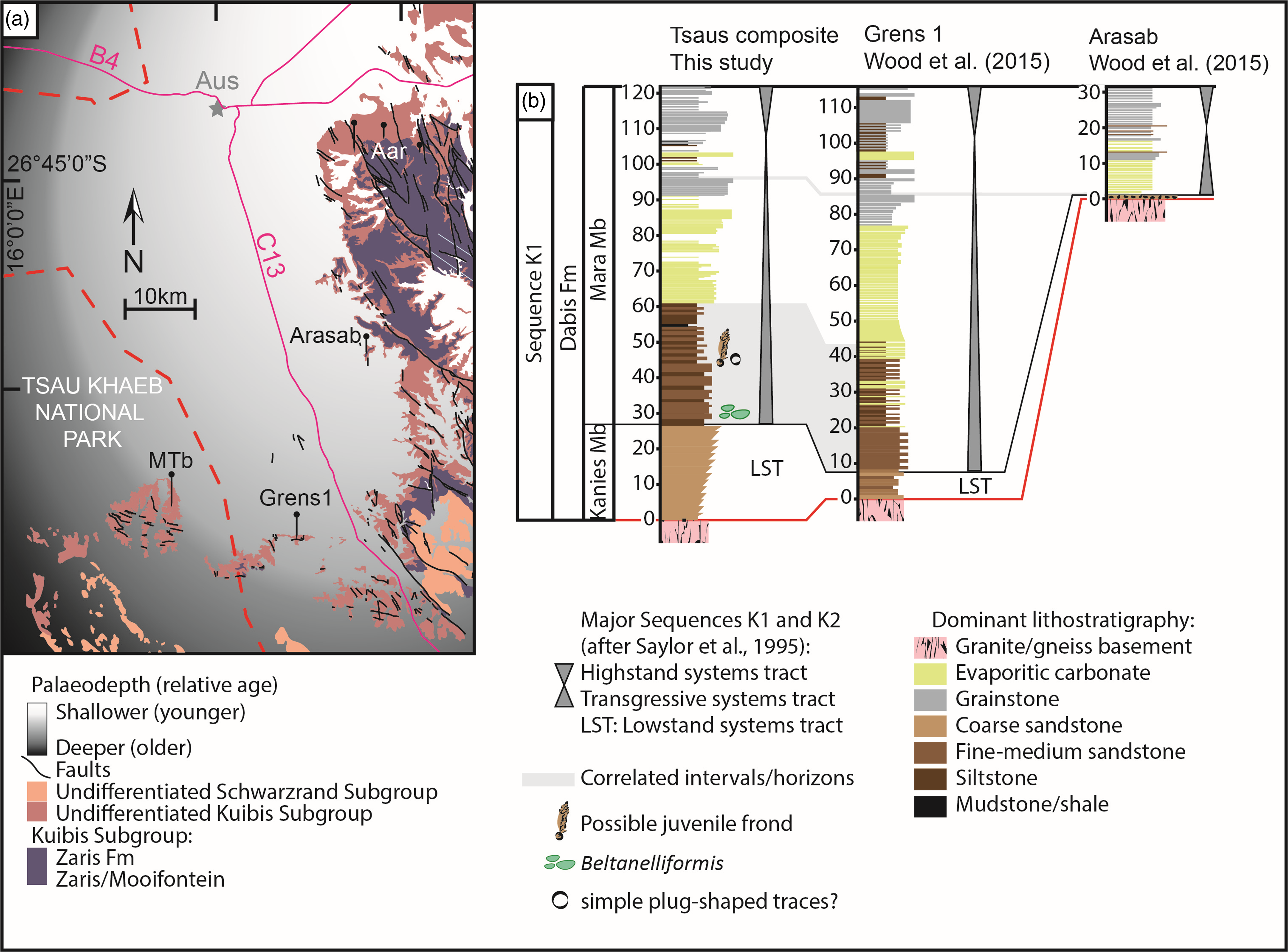
Figure 6. Lithostratigraphic and sequence stratigraphic correlation of the Kanies and lower Mara Members with documented fossil occurrences for sections deposited on the western edge of the Witputs Sub-basin, Tsaus Mountains, Grens 1 and Arasab. Inset map shows the positions of sections and relative palaeodepth (Modified from Bowyer et al. Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b).
The Kanies Mb has been previously described as consisting of monotonous planar-stratified and small-scale cross-stratified sandstones, and the Mara Member defined at its base by a transgressive surface commonly overlain by siltstone and the first appearance of carbonate interbeds (Saylor et al. Reference Saylor, Grotzinger and Germs1995). Carbonates that dominate the upper Mara Mb are thought to represent the remainder of the transgressive to highstand systems tract, ending in a sequence boundary (Saylor et al. Reference Saylor, Grotzinger and Germs1995). No clastic cycles have been previously recorded within the lower Mara Mb at other localities. We suggest here that the transgressive surface, which marks the start of the transgressive systems tract and the onset of basin-wide flooding (and the base of the Mara Mb), is most accurately marked in the section studied with the deposition of the first siltstones and fine sandstones, at ca. 27 m (Fig. 6). The underlying coarse sandstones of the Kanies Mb in the Tsaus Mountains are consistent with initial flooding to form a lowstand systems tract (Fig. 6).
We interpret the massive to planar-bedded sandstones from 27 to 45 m at the base of the lower Mara Mb to have been deposited by strong, sediment-laden currents, possibly accompanied by storms although no hummocky cross-stratification has been observed. The presence of asymmetric ripples indicates current flow, and units with symmetric and quasi-symmetric ripples may represent waves, currents or combined flows. Most possible biota in the lowermost Mara Mb are found in medium to fine-grained and rippled sandstones indicative of shallow, subtidal settings.
Other potentially contemporaneous localities globally contain macrofossil biotas, but in variable sedimentological settings. The Kanies and lower Mara members may pre-date deposition of the Khatyspyt Formation of the northern Siberian Platform, which has a maximum depositional age of ca. 550 Ma based on detrital zircon analyses of the underlying Maastakh Formation (Cherry et al. Reference Cherry, Gilleaudeau, Grazhdankin, Romaniello, Martoin and Kaufman2022). The Kanies and lower Mara members may also pre-date the Shibantan Mb of the middle Dengying Formation, South China, based on sedimentation rate estimates in the Nama succession, chemostratigraphic correlation between the lower Nama Group and Dengying Formation, and a zircon U-Pb age of 550.1 ± 0.6 Ma from an ash bed within the upper Doushantuo Fm (Mb IV), which immediately underlies the Dengying Formation (Yang et al. Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonaldn and Zhu2021).
The lower Zimnegory Formation of the eponymous White Sea area contains an ash bed recently re-dated at 553 ± 0.7 Ma (Yang et al. Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonaldn and Zhu2021), and so the shallow marine clastic deposits of the upper Zimnegory and lower Erga formations may immediately pre-date, or overlap with, deposition of the Kanies and lower Mara members. Lithofacies of the Zimnegory and lower Erga formations include lower-shoreface muds, middle-shoreface graded siltstone-shale couplets, inter-bedded prodelta sandstones and shales, and channelized sandstone beds of distributary-mouth bars (Grazhdankin, Reference Grazhdankin2004).
The Ediacara Mb of the Rawnsley Quartzite remains poorly constrained in time but likely contemporaneous with other White Sea assemblage localities >550 Ma and consists of a shallowing-upwards succession of up to five parasequences of shallow marine, nearshore to deltaic sandstones (Jenkins et al. Reference Jenkins, Ford and Gehling1983; Tarhan et al. Reference Tarhan, Droser, Gehling and Dzaugis2017; McMahon et al. Reference McMahon, Liu, Tindal and Kleinhans2020). This depositional environment is broadly similar to that of the Kanies and lower Mara members.
4.b. Affinity of fossil biota
Diverse structures of potentially biological origin are preserved. It is possible that some of the isolated raised, circular structures found on the rippled bedding plane of an in situ sandstone bed (Fig. 3a, top right) may represent holdfasts or trace fossils in the form of plug-shaped burrows. Similar structures, described from the Kliphoek Mb, have been assigned to Bergaueria (Darroch et al. Reference Darroch, Cribb, Buatois, Germs, Kenchington, Smith, Mocke, O’Neil, Schiffbauer, Maloney, Racicot, Turk, Gibson, Almond, Koester, Boag, Tweedt and Laflamme2021). The sharp edge of each structure, distinguishing them from the surrounding sediment, suggests that they are representative of sediment infill rather than sediment reconfiguration due to microbial activity, that is, microbially induced sedimentary structures (MISS) or fluid escape structures, which tend to display a domed morphology with sloped sides (Menon et al. Reference Menon, McIlroy, Liu and Brasier2016a). We note, however, that these examples are found on a bed top which is unusual for this ichnogenus. Further, a view of the base of the structure would be needed for confident assignment of these examples to Bergaueria. Indeed, some of these structures (Fig. 3a) could be partially filled spaces occupied by some form of tube or stalk. A view of the base of the structures and transverse sections will be required for more secure assignment of these examples.
The sinusoidal shape of the variably continuous chain of smaller raised, circular structures (Fig. 3c) indicates a configuration that is probably biologically mediated, rather than the result of geological processes. These structures appear similar to the morphology of the problematic body fossil Horodyskia, which displays a similar sigmoidal ‘string of beads’, and are known from 1.4 Ga to <557 Ma (Li et al. Reference Li, Chen, Pang, Tang, Wu, Yuan, Zhou and Xiao2023). The structure described herein is, however, systematically aligned on the rippled surface and so may have been orientated with respect to currents or topography. This may represent a mobile behavioural process that could be more suggestive of a trace fossil affinity. Alternatively, as the circular elements of Horodyskia tend to be more uniform and smaller, these circular structures might rather represent holdfast attachment points.
Possibly, the most notable specimen reported from the lower Mara Mb here is a potential frond, probably collapsed, preserved in direct association with a holdfast attachment (Fig. 3c). While the poor preservation of this specimen precludes confident taxonomic assignment, the general morphology is suggestive of a possible juvenile uniterminal rangeomorph or arboreomorph (Liu et al. Reference Liu, Kenchington and Mitchell2015).
A number of other raised, circular structures are found on the surfaces of in situ and float samples; however, these differ from those on the rippled bedding surface in that they are in far greater numbers and many appear to display a domed morphology with sloping sides (e.g. Fig. 3g). These often lack the sharp edges that characterize possible plug-shaped burrows. The specimen exhibiting large, closely spaced circular features (Fig. 3g) may be assigned, with a degree of confidence, to Nemiana which is synonymous with Beltanelliformis (Ivantsov, Reference Ivantsov2013). Beltanelliformis can occur in large numbers over extensive surface areas, but often cluster rather than being uniformly dispersed (Ivantsov et al. Reference Ivantsov, Gritsenko, Konstantinenko and Zakrevskaya2014). The affinity of Beltanelliformis has been the subject of considerable debate, with no consensus even as to a metazoan origin (e.g. Bobrovskiy et al. Reference Bobrovskiy, Hope, Ivantsov, Nettersheim, Hallmann and Brocks2018). Some more poorly preserved and less well-defined material that were attributed to these taxa have, in later studies, been found to be likely non-biogenic, such as fluid escape structures formed in the presence of microbial mats (Menon et al. Reference Menon, McIlroy, Liu and Brasier2016a). Poorly defined outlines may also be an indication of formation in a higher-energy environment. We also note that typical preservation of Beltanelliformis is as sandcasts on a bed sole but here our examples are, where known, on a bed top.
Where central depressions are evident (e.g. Fig. 3j), circular to oblong structures may represent Intrites, which was initially regarded as a trace fossil (e.g. Fedonkin, Reference Fedonkin1980; Crimes, Reference Crimes1987), but is now commonly regarded as a possible body fossil (e.g. Gehling et al. Reference Gehling, Narbonne and Anderson2000; McIlroy et al. Reference McIlroy, Crimes and Pauley2005; Jensen et al. Reference Jensen, Droser and Gehling2006), probably a palaeopascichnid (Kolesnikov and Desiatkin, Reference Kolesnikov and Desiatkin2022). Structures resembling Intrites have also been suggested to represent sedimentary surface textures or MISS associated with fluid escape under microbial mats (Davies et al. Reference Davies, Liu, Gibling and Miller2016; Menon et al. Reference Menon, McIlroy and Brasier2016b). Such structures are noted to be orientated in the direction of the prevailing current (Menon et al. Reference Menon, McIlroy and Brasier2016b), which may be consistent with a pronounced elongation present on some aligned individuals (e.g. Fig. 3d).
While some of the specimens presented herein may be assigned to Intrites, some individuals (Fig. 3j, insets) show central depressions that are characterized by similar shapes that, despite issues of preservation, appear more complex than a circular or oval depression. Rather than fluid escape, these may represent poorly preserved circular body fossils with elements of radial symmetry (e.g. pentaradialomorphs), many examples of which are found in the preceding White Sea Assemblage.
A sinuous, elongated, possibly ridged, structure is preserved on one float sample (Fig. 3k). Similar forms from the Nama Group have been identified as under-mat mining trace fossils, but the example here lacks the irregular, lobe-like extensions perpendicular on either side of the main sinuous form. While the lower Mara Mb does not represent the very shallow depositional setting where such forms have been found elsewhere in the Nama Group (Darroch et al. Reference Darroch, Cribb, Buatois, Germs, Kenchington, Smith, Mocke, O’Neil, Schiffbauer, Maloney, Racicot, Turk, Gibson, Almond, Koester, Boag, Tweedt and Laflamme2021), trace fossils interpreted as under-mat mining have been found in a wide range of depositional settings.
A number of float samples from the lower Mara Mb preserve long (>10 cm), raised sinusoidal structures (Fig. 4). We interpret such structures as likely sedimentary surface textures, formed either via microbial processes (e.g. MISS) or abiotic processes such as loading that may create morphologically similar features (see review of Davies et al. Reference Davies, Liu, Gibling and Miller2016), as they do not appear to disturb the underlying sediment, despite superficially resembling structures of biological origin due to their pattern and distribution. These may represent wrinkle structures that obscure the sedimentary surface known as ‘Kinneyia’ texture. ‘Kinneyia’ structures are mm-scale flat-topped, winding ridges and intervening troughs and pits, sometimes resembling small-scale interference ripples, that usually occur on upper surfaces of siltstone/sandstone beds, themselves frequently event deposits, which may have formed beneath microbial mats. These likely originated as thick microbial mats that were then further distorted by sedimentary loading, similar to features that have been noted elsewhere in the Nama Group (Noffke et al. Reference Noffke, Knoll and Grotzinger2002).
Microbial mats tend to be preserved as wrinkle structures on bedding surfaces in siliciclastic environments. Sediment pressure can create loading that forms moulds and casts at bedding planes, inducing the formation of wrinkle structures and complex ripple structures. Mat colonization is favoured by clean, fine-grained, quartz sands where hydrodynamic flow is sufficient to remove mud from mat surfaces but insufficient to erode biostabilized laminae, and preservation is favoured by subsequent burial without erosion (Noffke et al. Reference Noffke, Knoll and Grotzinger2002). Reduced hydrodynamic energy increases the thickness of the living mat layer (Noffke et al. Reference Noffke, Knoll and Grotzinger2002). But multi-directed ripples also commonly form in modern tidal environments without the presence of any microbial mats to produce morphologically indistinguishable structures to MISS, so any determination of MISS must be applied with care (Davies et al. Reference Davies, Liu, Gibling and Miller2016).
An array of lozenge-shaped structures that commonly form offset chains (Fig. 5) are superficially similar in morphology to treptichnids, a group of complex trace fossils (Jensen, Reference Jensen2003). Given the parallel alignment of many of these chains, however, and the fact that these specimens are considerably older (ca. >5 Myrs) than the current oldest documented treptichnids in the Nama Group (ca. 542.65 Ma; Darroch et al. Reference Darroch, Cribb, Buatois, Germs, Kenchington, Smith, Mocke, O’Neil, Schiffbauer, Maloney, Racicot, Turk, Gibson, Almond, Koester, Boag, Tweedt and Laflamme2021; Nelson et al. Reference Nelson, Ramezani, Almond, Darroch, Taylor, Brenner, Furey, Turner and Smith2022), it is unlikely that they represent treptichnids. Instead, such features probably formed due to the infill of aligned, shrinkage or en-echelon cracks (cf. Glaessner, Reference Glaessner1969). The similarity of these probable pseudofossils to trace fossils demonstrates the difficulty in ascribing a biotic origin and taxonomic designation to many structures in sedimentary units.
4.c. Stratigraphic significance of the biota
The Verkhovka, Zimnegory and Erga (Yorga) formations of the White Sea area contain a high diversity of soft-bodied fossils corresponding to the White Sea Assemblage (e.g. Grazhdankin, Reference Grazhdankin2004; Fedonkin et al. Reference Fedonkin, Vickers-Rich, Swalla, Trusler and Hall2012). These include dickinsoniids, triradialomorphs and bilateralomorphs (e.g. Parvancorina) (Fedonkin et al. Reference Fedonkin, Vickers-Rich, Swalla, Trusler and Hall2012). Deposits of the Ediacara Mb of the Rawnsley Quartzite preserve soft-bodied macrofossils and trace fossils in an abundance and diversity similar to those of the White Sea area, but not yet observed in the Kanies or Mara members (e.g. Droser et al. Reference Droser, Gehling, Tarhan, Evans, Hall, Hughes, Hughes, Dzaugis, Dzaugis, Dzaugis and Rice2019). We may assume that the Kanies and lower Mara members postdate deposition of the Ediacara Mb, South Australia and the Zimnegory and Erga formations of the White Sea area based on the absence of key marker body fossils (e.g. Kimberella, Parvancorina, Eoandromeda and Tribrachidium) from the entire Nama Group succession.
The potential body fossils (Beltanelliformis and possible juvenile rangeomorph/arboreomorph frond) and plug-shaped burrows (?Bergaueria) noted from the lower Mara Mb have been reported from strata in other regions that pre-date deposition of the lower Mara Mb and have also been reported from strata that postdate deposition of the Mara Mb in the Nama Group succession and/or other temporally correlative depositional settings. Taxa such as Aspidella and Rangea are also known from both the older White Sea assemblage-aged strata elsewhere and from overlying strata within the Nama Group succession, so their absence from the Kanies and lower Mara members at Tsaus is not noteworthy. Indeed, no fossils uniquely characteristic of either the White Sea or Nama assemblages have yet been found in the Kanies or lower Mara members.
Most samples studied in the Tsaus Mountains to date represent either in situ bed tops or float samples of uncertain orientation. Upper bedding surfaces will preferentially record trace fossils and holdfasts, rather than the internal or soles of storm event beds which typically contain abundant entrained body fossils (e.g. Ernietta) in the Nama Group (e.g. Narbonne et al. Reference Narbonne, Saylor and Grotzinger1997). Indeed in the Nama assemblage, the typical preservation of trace fossils is on bed soles, not bed tops and likewise mainly bed sole surfaces of bedding planes and their counterparts preserve the White Sea Assemblage fossils of the Ediacara Mb (e.g. Tarhan et al. Reference Tarhan, Droser, Gehling and Dzaugis2017; Droser et al. Reference Droser, Gehling, Tarhan, Evans, Hall, Hughes, Hughes, Dzaugis, Dzaugis, Dzaugis and Rice2019). So the apparent absence of typical White Sea Assemblage fossils from the Kanies and lower Mara members may, therefore, not be significant. Indeed, our sampling is very preliminary and undertaken without any excavation and this is likely to have limited our discoveries, in particular possible biota preserved on bed soles (see ‘Pledge’ below). Indeed, if the studied outcrops of the Tsaus Mountains had provided accessible and clean sandstone bed soles without the need for destructive excavation, then their scrutiny may well have resulted in a very different and more diverse tally of fossils.
5. Redefinition of the Nama assemblage
Evolutionary faunas are usually defined based on groups of high-level taxa that typically appear around the same time and display a broadly synchronous logarithmic increase in biodiversity followed by extinctions, with the increase in diversity associated with successive faunas coinciding with a decline of the previously dominant fauna (Sepkoski, Reference Sepkoski1981). The proposed assemblages/palaeocommunities of the Ediacaran, unusually, have a temporal component. This is particularly notable with the Nama Assemblage, where all taxa present between ca. 550 Ma and <538 Ma are ascribed to that assemblage irrespective of morphology or taxonomic grouping.
A representative compilation of Ediacaran to earliest Cambrian macrofossil and ichnofossil morphogroups and genera (ca. 580–528 Ma), calibrated with available radiometric dates and to a composite δ13C curve (updated after Bowyer et al. Reference Bowyer, Zhuravlev, Wood, Shields, Zhou, Curtis, Poulton, Condon, Yang and Zhu2022, Reference Bowyer, Zhuravlev, Wood, Zhao, Sukhov, Alexander, Poulton and Zhu2023a, Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b; Nelson et al. Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023), reveals some notable trends (Fig. 7). As the inter-relationships of proposed Ediacaran morphogroups are so poorly understood, here the ranges of genera become important.
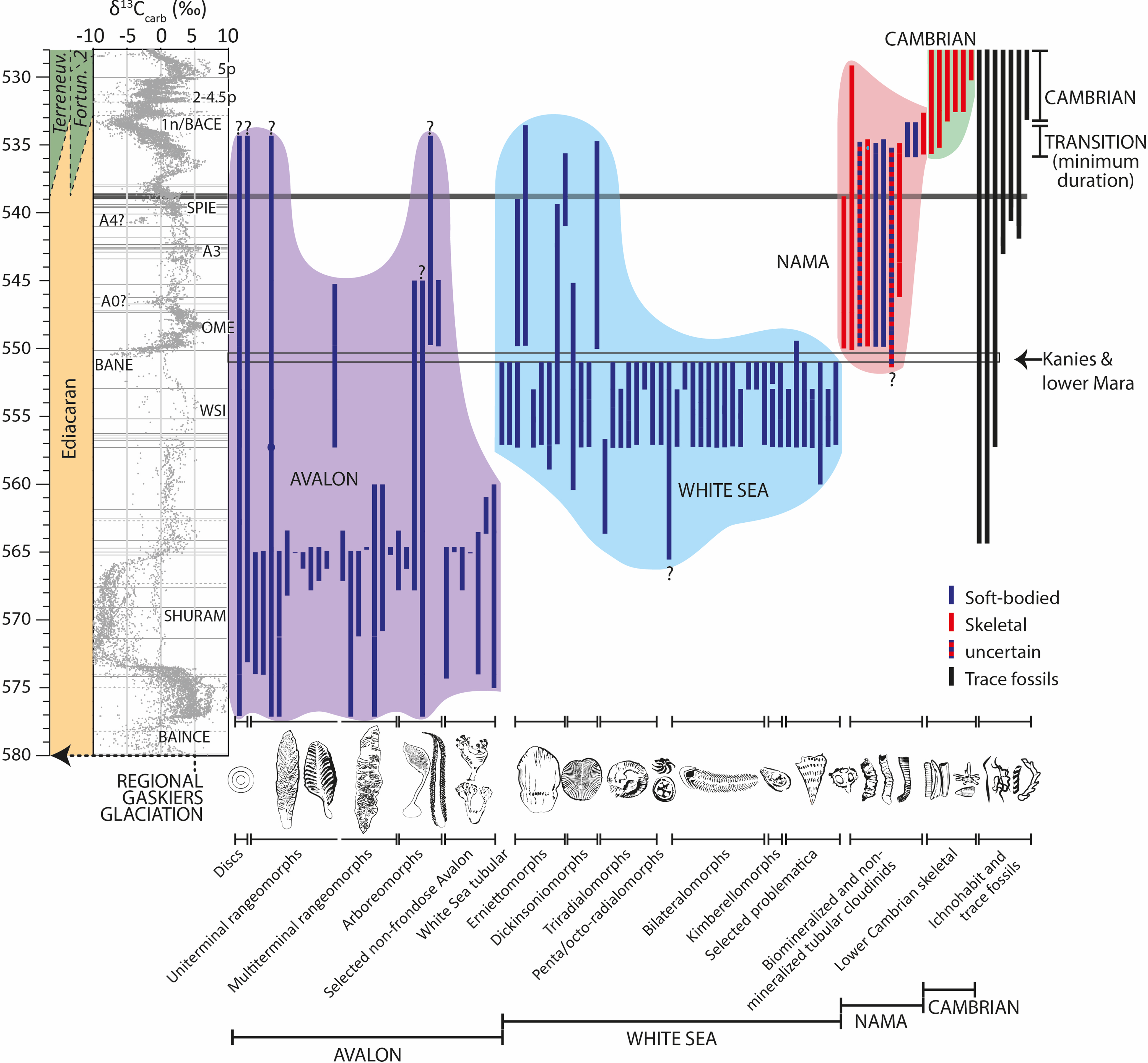
Figure 7. Compilation of ranges of known major Ediacaran to earliest Cambrian macrofossils (soft-bodied and skeletal) and major trace fossil groups (ca. 580–528 Ma) calibrated to radiometric dates and an updated composite δ13C curve (after Yang et al. Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonaldn and Zhu2021; Bowyer et al. Reference Bowyer, Zhuravlev, Wood, Shields, Zhou, Curtis, Poulton, Condon, Yang and Zhu2022, Reference Bowyer, Zhuravlev, Wood, Zhao, Sukhov, Alexander, Poulton and Zhu2023a, Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b; Nelson et al. Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023). Figure shows the possible age of the Kanies and lower Mara Members based on the most parsimonious global radiometric, chemostratigraphic and preliminary biostratigraphic considerations. Proposed re-definition of Avalon, White Sea and Nama assemblages is shown, based on morphogroup membership alone, with the appearance of the transitional and Cambrian assemblage from ca. 535 to 536 Ma. Question marks denote uncertainty in the precise age of specific formations/members (see Supplementary Information of Bowyer et al. Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b).
The Avalon Assemblage appears ca. ≥575 Ma and is entirely soft-bodied, consisting of discs, uniterminal and multiterminal rangeomorphs, arboreomorphs and other non-frondose taxa (e.g. Matthews et al. Reference Matthews, Liu, Yang, McIlroy, Levell and Condon2020). There is an apparent decline in biodiversity occurring ca. 565–560 Ma, but it remains unclear as to whether this is associated with sampling and/or taphonomic biases. Regardless, several taxa of the Avalon Assemblage morphogroups persist, for example, the disc Hiemalora, rangeomorph Charnia and arboreomorph Arborea until at least ca. 545 Ma, and ‘holdfasts’ until at least ca. 538 Ma. A new rangeomorph taxon, Rangea, appears at ca. 557.5 Ma and persists until ca. 545 Ma, and at least one new arboreomorph taxon, Paracharnia, appears <550 Ma in South China and persists until at least ca. 545 Ma. Following definitions used to determine Phanerozoic evolutionary faunas, all such taxa that belong to pre-existing morphogroups, even new genera, can be considered to be members of the Avalon Assemblage even though they occur during the Nama assemblage time interval.
Most White Sea assemblage taxa appear from ca. 557.5 to 555 Ma and consist of entirely new morphological groups (e.g. erniettomorphs, dickinsoniomorphs, triradialomorphs, pentaradialomorphs, tetraradialomorphs, bilateralomorphs, kimberellomorphs and some other problematic forms). This assemblage is currently supposed to have largely disappeared by ca. 550 Ma, coincident with a marked decline in biodiversity (Grazhdankin, Reference Grazhdankin2014; Yang et al. Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonaldn and Zhu2021; Evans et al. Reference Evans, Tu, Rizzo, Surprenant, Boan, McCandless, Marshall, Xiao and Droser2022). The updated compilation supports a dramatic drop in diversity ca. 551 Ma, but some erniettomorph taxa persist until at least ca. 539 Ma, such as Pteridinium and Nasepia. New and distinctive erniettomorph taxa also appear, probably Ernietta and Swartpuntia at ca. 550 Ma and ca. 541 Ma, respectively, which persist until ca. 534 Ma and ca. 535.5 Ma, respectively (e.g. Nelson et al. Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023). A partial specimen assigned to Dickinsonia has also been reported from the Shibantan Member of the Dengying Formation (<550 Ma) (Wang et al. Reference Wang, Chen, Pang, Zhou, Xiao, Wan and Yuan2021). By the same reasoning, notwithstanding the appearance of new taxa in these morphogroups, all erniettomorphs and dickinsoniomorphs can be considered to be members of the White Sea Assemblage.
The fossil assemblage of the lower Mara Mb likely immediately postdates or is contemporaneous with the major loss of White Sea biodiversity (Fig. 7). Remaining White Sea morphogroups, the erniettomorphs and dickinsoniomorphs persist, and from ca. 550 Ma onwards, they are joined by new and diverse calcified tubular and non-tubular metazoans (e.g. Cloudina, Namacalathus, Sinotubulites and Corumbella), non-calcified tubular metazoans (e.g. Conotubus and Gaojiashania), and more complex trace fossils (e.g. Torrowangea and Streptichnus) (Fig. 7).
As currently defined, the Nama Assemblage consists of a low diversity of soft-bodied taxa (largely erniettomorphs with subordinate rangeomorphs, arboreomorphs, and non-frondose soft-bodied forms) that are present in strata <550 Ma, but we argue these are representatives of morphological groups that appeared in older strata within the preceding Avalon and White Sea assemblages. By contrast, soft and calcified tubular metazoans (broadly the ‘cloudinids’ to distinguish them from preceding non-cloudinid tubular forms such as Aulozoon and Funisia; for example, Droser et al. Reference Droser, Gehling, Tarhan, Evans, Hall, Hughes, Hughes, Dzaugis, Dzaugis, Dzaugis and Rice2019) as well as other skeletal taxa represent the appearance of taxa that belong to new morphogroups that postdate the White Sea Assemblage interval. The proposed Terminal Ediacaran biozone, which is considered to correspond to the Nama palaeocommunity (Muscente et al. Reference Muscente, Bykova, Boag, Buatois, Mángano, Eleish, Prabhu, Pan, Meyer, Schiffbauer, Fox, Hazen and Knoll2019), includes both soft-bodied (such as Ernietta and Swartpuntia) and novel soft and calcified tubular metazoans. While we agree with both the validity and utility of this biozone, clarification of the separate evolutionary assemblage concept deems that we exclude the soft-bodied erniettomorphs, rangeomorphs and arboreomorphs from the Nama Assemblage. Indeed, network analysis confirms that soft and calcified tubular metazoans together with metazoan trace fossils, but not the soft-bodied erniettomorphs, rangeomorphs and arboreomorphs, are the best connected taxa in the Terminal Ediacaran biozone (Muscente et al. Reference Muscente, Bykova, Boag, Buatois, Mángano, Eleish, Prabhu, Pan, Meyer, Schiffbauer, Fox, Hazen and Knoll2019). This distinction is supported by the observation that holdover Avalon and White Sea taxa occupied different environmental settings from soft and calcified tubular and other biomineralized metazoans, precluding direct competition for resources (Wood et al. Reference Wood, Liu, Bowyer, Wilby, Dunn, Kenchington, Hoyal Cuthill, Mitchell and Penny2019). This underscores the definition of the Avalon, White Sea and Nama assemblages as evolutionary faunas.
We hence propose re-definition of the Nama Assemblage to be based on new morphogroups alone – calcified metazoans (including cloudinids), soft-bodied tubular cloudinids, and later complex trace fossils – with their first appearance ca. 550 Ma and persistence until ca. 539–534 Ma (Fig. 7). A new Cambrian assemblage appears from ca. 536 Ma (updated after Bowyer et al. Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b; Nelson et al. Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023), and overlaps in time with cloudinids of the Nama assemblage (e.g. Topper et al. Reference Topper, Betts, Dorjnamjaa, Li, Li, Altanshagai, Enkhbaatar and Skovsted2022).
6. Conclusions
We describe a thick succession of the Kanies and lower Mara members from a new exposure in the Tsaus Mountains of southwestern Namibia. Here, the Kanies Mb records lowstand clastic deposition atop the metamorphosed Proterozoic basement, and the overlying lower Mara Mb records the onset of basin-wide marine transgression. While no biota has been previously reported from the Kanies or lower Mara members, we describe structures that may represent body (Beltanelliformis and a possible juvenile rangeomorph/arboreomorph frond) and trace (?Bergaueria) fossils, in addition to sedimentary surface textures from the lower Mara Mb.
While the lower Mara Mb does not appear to preserve a biotic diversity comparable to potentially contemporaneous successions, evidence of trace and/or body fossils in the oldest Nama Group is essential to a complete understanding of biotic evolution during the late Ediacaran. The taxa noted here represent holdover taxa from the soft-bodied and trace fossil White Sea assemblage that appear prior to the appearance of diverse tubular and biomineralized metazoans. Our findings and fossil inventory are, however, preliminary and biased by sampling of only float and exposed bed tops given the need for non-destructive excavation that precluded exploration of potential bed sole surfaces.
Based on a new compilation of major Ediacaran-transitional Cambrian taxa and morphogroups within an updated global age model (Yang et al. Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonaldn and Zhu2021; Bowyer et al. Reference Bowyer, Zhuravlev, Wood, Shields, Zhou, Curtis, Poulton, Condon, Yang and Zhu2022, Reference Bowyer, Uahengo, Kaputuaza, Ndeunyema, Yilales, Alexander, Curtis and Wood2023b; Nelson et al. Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023), we propose re-definition of the Nama Assemblage based on the appearance of new taxa of calcified metazoans, soft-bodied and calcified tubular cloudinids, as well as later complex trace fossils, which first appear globally ca. 550 Ma, postdating deposition of the lower Mara Mb, and persist until at least 539 Ma.
Other Nama Group sites rich in potential soft-bodied fossils have been extensively excavated. The Tsaus Mountains are a pristine and unique environment, unspoilt by direct human action. We suggest that all who visit the site should be able to make a pledge in every associated publication, which we make below, and will leave the area and geology undisturbed. A similar approach might be adopted for other pristine sites.
7. Pledge
The area studied in this manuscript is a pristine nature reserve. Following World Heritage Convention, all sampling for this study was undertaken without destructive excavation of in situ beds, as a requirement of access and sampling. The environment was left visibly as found on arrival.
Data accessibility
All datasets used for the analyses are available in the main text.
Acknowledgements
We thank Harry Tjihukununa and Directorate of Wildlife and National Parks for permission to enter the (Tsaus Mountains) site within the Tsau/Khaeb National Park and are indebted to ranger Alex Mowe from the Directorate of Wildlife and National Parks, and Helke Mocke from the Geological Survey of Namibia, for their help and support. We are grateful for funding from UKRI NERC Project NE/T008458/1 (R.W. and F.B.). We thank Soren Jensen and a further anonymous reviewer whose reviews substantially improved this manuscript. For the purpose of open access, the authors have applied a Creative Commons Attribution (CC BY) licence to any Author Accepted Manuscript version arising.
Authors’ contributions
All authors conducted fieldwork and conceptualization of the research. R.W, R.A., M.Y. and F.B. – original draft and writing. F.B. and M.Y. – figures. R.W. – funding acquisition. R.W., C.-I.U. and A.C. – supervision. All authors – review and editing.
All authors gave final approval for publication and agreed to be held accountable for the work performed therein.
Financial support
This study was funded by UKRI NERC Project NE/T008458/1 to RW.
Competing interests
We declare we have no competing interests.








 Open access
Open access