


Introduction
Though now restricted to southern landmasses, lungfishes were present in North America for much of the Paleozoic and Mesozoic, dating back to the Devonian Period. Early representatives display broad diversity in skull and dental morphology, suggesting differences in trophic adaptations and representation of multiple clades (e.g., Long, Reference Long2010; Pardo et al., Reference Pardo, Huttenlocker, Small and Gorman2010). Post-Triassic dipnoans of North America, however, are represented almost entirely by small samples (or unique occurrences) of isolated tooth plates—a fossil record that is poorly suited to detailed phylogenetic or paleobiologic understanding, and that is therefore permissive of contrasting perspectives. For example, Martin (Reference Martin1982) proposed family-level distinction of the two major morphs (flat and sharp-crested, referred to Ceratodontidae and Ptychoceratodontidae, respectively) seen among North American Jurassic–Cretaceous varieties. Alternate hypotheses of relationships have been suggested, however (e.g., Kirkland, Reference Kirkland1987, Reference Kirkland1998), and in view of the fact that the tooth plates are broadly similar in gross anatomical and histological features, they are generally referred to Ceratodus (Agassiz, Reference Agassiz1838; see diagnosis in Kemp, Reference Kemp1993), which has long been regarded as a form genus (Schultze, Reference Schultze1981; Kirkland, Reference Kirkland1987). That this fossil record imperfectly reflects taxonomic diversity is illustrated by cranial remains recently described for one species (named Ceratodus guentheri by Marsh, Reference Marsh1878; and compared favorably to Ptychoceratodus by Martin, Reference Martin1982 and some subsequent workers) from the Morrison Formation, which is distinctive enough to warrant placement in a separate genus, Potamoceratodus (Pardo et al., Reference Pardo, Huttenlocker, Small and Gorman2010). On the other hand, tooth plates of the closest living species, Neoceratodus forsteri, are sufficiently variable (particularly according to growth stage) that fossils dating as far back as the Early Cretaceous are attributable to it (Kemp and Molnar, Reference Kemp and Molnar1981; see also Kemp, Reference Kemp1997). The opposing issues of general morphologic conservatism and intraspecific variability pose challenges to interpreting a fossil record that consists mainly of rare, widely scattered occurrences of isolated tooth plates.
Yet the record of North American Jurassic–Cretaceous lungfish is informative in spite of its imperfection. Recognizable species-level distinctions can often be made between isolated specimens or small samples of Ceratodus tooth plates (see Kemp, Reference Kemp1993 and references therein; Kirkland, Reference Kirkland1987, Reference Kirkland1998). Moreover, the contrasting morphology between the major types of tooth plates (flat versus high-crested) suggests diversity in dietary preference. Species of Ceratodus (and Potamoceratodus) are inferred to have been faunivorous to omnivorous; low-crowned, flat tooth plates are interpreted to reflect adaptation for durophagy, suggesting more reliance on mollusks and, perhaps, vertebrates (Kirkland, Reference Kirkland1987; Bakker, Reference Bakker2009; Shimada and Kirkland, Reference Shimada and Kirkland2011). It is also noteworthy that some of the flat-crowned (and presumably durophagous) taxa were the largest fishes of their respective communities; one unnamed species attained an estimated length of ~4 m (Shimada and Kirkland, Reference Shimada and Kirkland2011). Smaller species with high-crested tooth plates—suggestive of shearing function—may have relied more heavily on fare having negligible mineralized tissue, such as soft-bodied invertebrates (Parris et al., Reference Parris, Grandstaff and Banks2007). Dietary preference also presumably varied ontogenetically, as in the living Australian lungfish (Neoceratodus forsteri), whose dentition progresses from sharp cones, suitable mainly for apprehending food items, to well-worn crushing plates, which are capable of processing mollusks and other hard-bodied prey (Kemp, Reference Kemp1986).
Lungfishes were geographically and stratigraphically widespread in the Cretaceous of North America, and include fossils ranging from the Albian of Montana and Wyoming (Ostrom, Reference Ostrom1970; Oreska et al., Reference Oreska, Carrano and Dzikiewicz2013) to the Late Cretaceous (probably Campanian) of New Jersey (Parris et al., Reference Parris, Grandstaff and Gallagher2004). Occurrences within this broad temporal and geographic range are sparse and rare, however. All but two are from western North America (Parris et al., Reference Parris, Grandstaff and Gallagher2004; Frederickson et al., Reference Frederickson, Lipka and Cifelli2016), and overall taxonomic representation is meager. Herein we report new dipnoan fossils from Cretaceous units spanning the Valanginian–Cenomanian of North America’s Western Interior Basin (see Kauffman and Caldwell, Reference Kauffman and Caldwell1993). These include representation of four new species named herein: Ceratodus kempae n. sp., C. kirklandi n. sp., C. molossus n. sp., and C. nirumbee n. sp. We also add to knowledge of Jurassic C. stewarti Milner and Kirkland, Reference Milner and Kirkland2006 by referring an upper tooth plate to the species. These new fossils, together with critical appraisal of other occurrences, provide basis for evaluating ceratodontid diversity (morphologic and taxonomic) through the Cretaceous of North America.
Materials and methods
Standardized angles and reference points (Kirkland, Reference Kirkland1987 and references therein; abbreviations for structures used as basis for measuring angles are illustrated in Fig. 1) were measured from photographs of specimens described herein (Table 1). We present these data mainly to provide consistent frame of reference with respect to previous studies on North American ceratodontids (e.g., Kirkland, Reference Kirkland1987, Reference Kirkland1998; Parris et al., Reference Parris, Grandstaff and Gallagher2004, Reference Parris, Grandstaff and Banks2014; Main, Reference Main2013; Main et al., Reference Main, Parris, Grandstaff and Carter2014). Phylogenetic utility of these measurements is limited, largely owing to a lack of basis for assessing variability: most North American taxa are known by one or a few specimens. Further, in Australian lungfishes, angles of the ridge crests can be very similar, even in visually distinct tooth plates (Kemp and Molnar, Reference Kemp and Molnar1981; Kemp, Reference Kemp1997). In this study, we compared gross morphology using the published descriptions and anatomical terminology of Kirkland (Reference Kirkland1987). Since there is currently no single set of characters that can be used to accurately diagnose all fossil lungfish species, qualitative approaches that follow the outlines suggested by Kirkland (Reference Kirkland1987) and Kemp (Reference Kemp1997) are used to define the taxonomic boundaries between species. Finally, no attempt is made to reconstruct phylogenetic relationships, because most of these specimens are composed solely of tooth plates having limited morphological information.

Figure 1 A hypothetical Ceratodus right prearticular plate showing orientation, basis for angles, and abbreviations for crests, following Kirkland (Reference Kirkland1987, Reference Kirkland1998).
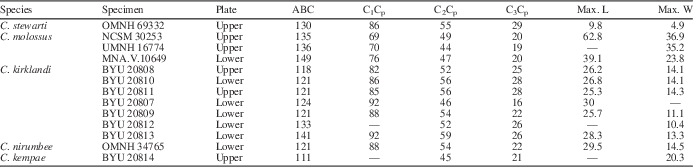
Table 1 Measurements of Ceratodus tooth plates described in this work. Angles, in degrees, were measured following protocol outlined and illustrated by Kirkland (Reference Kirkland1987, fig. 1; see Fig. 1 herein). ABC=angle between mesial (line A–B) and lingual (line B–C) borders, C1Cp=angle between lines defined by first (C1) and last (Cp) crests, C2Cp=angle between lines formed by second (C2) and last (Cp) crests, C3Cp=angle between lines formed by third (C3) and last (Cp) crests, Max. L=maximum length is the end of C1 to Cp from the labial margin in mm, Max. W=maximum width is the labial margin to the lingual-mesial margin junction in mm.
Except where indicated otherwise, specimens reported herein were collected from Federal lands administered by the U.S. Bureau of Land Management, which maintains and restricts access to specific locality coordinates. For southern and central Utah, the rock unit traditionally cited as the Dakota Formation is herein termed the Naturita Formation (see Young, Reference Young1960; Carpenter, Reference Carpenter2014).
Repositories and institutional abbreviations
Specimens mentioned in this study are deposited in the following institutions: AMNH, American Museum of Natural History, New York, New York, U.S.A.; BYU, Paleontological Collections, Brigham Young University, Provo, Utah, U.S.A.; DMNS, Denver Museum of Nature and Science, Denver, Colorado, U.S.A.; KUVP, Vertebrate Paleontology Collection, University of Kansas Natural History Museum, Lawrence, Kansas, U.S.A.; MCZ, Museum of Comparative Anatomy, Harvard University, Cambridge, Massachusetts; MNA, Museum of Northern Arizona, Flagstaff, Arizona, U.S.A.; MOR, Museum of the Rockies, Bozeman, Montana; NCSM, North Carolina Museum of Natural Sciences, Raleigh, North Carolina, U.S.A.; NJSM, New Jersey State Museum, Trenton, New Jersey, U.S.A.; OMNH, Sam Noble Oklahoma Museum of Natural History, Norman, Oklahoma, U.S.A.; SDSM, Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.; TATE, Tate Museum, Casper, Wyoming, U.S.A.; UCM, University of Colorado Museum, Boulder, Colorado, U.S.A.; UMNH-VP, Vertebrate Paleontology Collection, Utah Museum of Natural History, Salt Lake City, Utah, U.S.A.; UNSM, University of Nebraska State Museum, Lincoln, Nebraska, U.S.A.; USNM, National Museum of Natural History (formerly United States National Museum), Smithsonian Institution, Washington, D.C., U.S.A.; UTA-AAS, University of Texas at Arlington, Arlington Archosaur Site, Arlington, Texas, U.S.A. (numbers cited from Main et al., Reference Main, Parris, Grandstaff and Carter2014, specimens are now deposited at the Perot Museum of Science and Nature, Dallas, Texas, U.S.A.); UWM, University of Wyoming Museum, Laramie, Wyoming, U.S.A.; VOF, Valley of Fire State Park, currently curated at the Nevada State Museum and Historical Society, Las Vegas, Nevada, U.S.A; YPM, Yale Peabody Museum, New Haven, Connecticut, U.S.A.
Systematic paleontology
Subclass Dipnoi Müller, Reference Müller1846
Order Ceratodontiformes Berg, Reference Berg1940
Family Ceratodontidae Gill, Reference Gill1872
Genus Ceratodus Agassiz, Reference Agassiz1838
Type species
Ceratodus latissimus Agassiz, Reference Agassiz1838.
Ceratodus stewarti Milner and Kirkland, Reference Milner and Kirkland2006

Figure 2 Ceratodus from North America. (1) C. stewarti, OMNH 69332, left pterygopalatine plate in occlusal view; OMNH locality V1737, Emery County, Utah, Kayenta Formation, Lower Jurassic (Sinemurian). (2–7) C. molossus n. sp. (2, 3) Occlusal and dorsal views, respectively, of NCSM 30253 (holotype), right pterygopalatine plate; locality near Emery, Emery County, Utah, Mussentuchit Member, Cedar Mountain Formation, Upper Cretaceous (Cenomanian). (4, 5) Occlusal and dorsal views, respectively, of UMNH 16774, right pterygopalatine plate; locality near Escalante, Garfield County, Utah, Naturita Formation, Upper Cretaceous (Cenomanian). (6, 7) Occlusal and inferior views, respectively, of MNA.V.10649, right prearticular plate; MNA locality 1067, Garfield County, Utah, Naturita Formation, Upper Cretaceous (Cenomanian). (8, 9) C. nirumbee n. sp., occlusal and inferior views, respectively, of OMNH 34765 (holotype), right prearticular plate; OMNH locality V1061, Carbon County, Montana, Cloverly Formation, Lower Cretaceous (Albian). (10, 11) C. kempae n. sp., BYU 20814 (holotype), right pterygopalatine plate in occlusal and dorsal views, respectively; pter.=pterygoid; BYU locality 1783, Grand County, Utah; Yellow Cat Member, Cedar Mountain Formation, Lower Cretaceous (Valanginian). (12, 13) C. frazieri, OMNH 60408, right prearticular plate in occlusal and inferior views; OMNH locality 1075, Big Horn County, Montana; Cloverly Formation, Lower Cretaceous (Albian). (14, 15) C. gustasoni, OMNH 66043, left prearticular plate in occlusal and inferior views, respectively; OMNH locality V808, Garfield County, Utah; Naturita Formation, Upper Cretaceous (Cenomanian).
2006 Ceratodus stewarti Reference Milner and KirklandMilner and Kirkland, p. 515, fig. 7B.
Holotype
Prearticular plate with fused prearticular (UMNH-VP 16037), Early Jurassic, from the Whitmore Point Member, Moenave Formation; 12 km east of St. George, Washington County, Utah, U.S.A. (Milner and Kirkland, Reference Milner and Kirkland2006, fig. 1B).
Emended diagnosis
Lower tooth plates with four primary ridges (the fourth ridge being twinned); upper tooth plate has five ridges with the fifth being twinned; both upper and lower tooth plates have relatively tall ridges, with C1Cp angle being slightly acute.
Occurrence
Lower Jurassic (Hettangian–Sinemurian), southern Utah. The newly referred specimen (OMNH 69332) is from OMNH locality V1737, 4 km west of Goblin Valley State Park, Emery County, Utah, U.S.A.; Kayenta Formation (Lower Jurassic, Sinemurian, see Lucas et al., Reference Lucas, Heckert and Tanner2005).
Description
Milner and Kirkland (Reference Milner and Kirkland2006) described C. stewarti based on a single prearticular tooth plate. The prearticular plate of this species is distinct in its possession of a twinned fourth ridge (here defined as a single crest arising from central mound that splits into two crests before reaching the labial margin), such that five functional cutting surfaces are present; a trait not known to occur in any other ceratodontid from the Jurassic. This species is most similar to Potamoceratodus guentheri in the possession of tall ridges, with an acute C1Cp angle. The upper (pterygopalatine) plate attributed to the species here (Fig. 2.1) similarly has a twinned fifth ridge crest, giving this specimen six functional cutting surfaces. The upper plate, like the lower, has a slightly acute C1Cp angle and well-defined crests or ridges.
Materials
UMNH-VP 16037 (holotype), prearticular plate with fused prearticular; and OMNH 69332, left pterygopalatine tooth plate.
Remarks
We have emended the diagnosis of C. stewarti in order to account for the referred pterygopalatine plate, the morphology of which was anticipated by Milner and Kirkland (Reference Milner and Kirkland2006). The referred specimen extends the geologic range of C. stewarti upward to the Sinemurian. Milner and Kirkland (Reference Milner and Kirkland2006) also mentioned a prearticular tooth plate, MCZ 13865, from the Kayenta Formation of nearby northern Arizona (see Lucas et al., Reference Lucas, Heckert and Tanner2005), referring it to Ceratodus cf. C. guentheri. We agree with this assignment (see discussion).
Ceratodus kirklandi new species
Figure 3.1–3.9, Supplemental Data 1, 2, Table 1

Figure 3 Ceratodus kirklandi n. sp. All specimens are from BYU locality 1667, Grand County, Utah; Yellow Cat Member, Cedar Mountain Formation, Lower Cretaceous (Valanginian). (1, 2) BYU 20808 (holotype), right pterygopalatine plate in occlusal and dorsal views, respectively; pter.=pterygoid. (3) BYU 20811, left pterygopalatine plate in occlusal view. (4, 5) BYU 20809, left prearticular plate in occlusal and inferior views, respectively. (6) BYU 20807, right prearticular plate in occlusal view. (7) BYU 20812, left prearticular plate in occlusal view. (8) BYU 20813, left prearticular plate in occlusal view. (9) BYU 20810, left prearticular plate in occlusal view.
Holotype
BYU 20808, right pterygopalatine plate with attached pterygopalatine, Early Cretaceous (Valanginian, based on Hendrix et al., Reference Hendrix, Moeller, Ludvigson, Joeckel and Kirkland2015), lowest part of Yellow Cat Member, Cedar Mountain Formation; BYU locality 1667, 14 km southwest of Cisco, Grand County, Utah, U.S.A.
Diagnosis
A ceratodontid of moderate size; pterygopalatine plate with five ridges and a fifth ridge crest that is variably expanded into a flat crushing surface, with twinning present. Prearticular plate with four main ridges; fourth ridge always twinned, giving the specimens five ridge crests. Prearticular plates have a groove on the lingual edge of the occlusal surface to accommodate the inner angle of the upper tooth plate.
Occurrence
Known only from the type locality, Grand County, Utah; Yellow Cat Member of the Cedar Mountain Formation (Lower Cretaceous).
Description
The available sample of this species includes two pterygopalatine and five prearticular tooth plates, all with plate-bearing element attached. Pterygopalatine plates are similar in size and complete prearticular plates vary only slightly in maximum length (Table 1). All plates are sharply crested and angles between crests are relatively acute (see C1Cp values, Table 1). The occlusal surfaces on the margins of both the upper and lower plates show wear from the opposing tooth plates. In both plates the wear is angled, sharpening the ridges and crests with increased use. Wear on the lower plates is more heavily developed on the mesial side of the ridge crests, while on the upper plates it is heavier on the lingual side of the ridge crests. In general it appears that the pterygopalatine plates are more deeply incised than the prearticular plates.
The pterygopalatine plates have five main crests, each bearing up to eight denticles (C2 of BYU 20808). Crest five of BYU 20808 broadens into a flat surface distally and a faint sixth crest is present at the posterior margin (Fig. 3.1). The pterygopalatine (the bone to which the plate is fused) is undiagnostic, except for the pterygoid process, which is relatively small and pointed by comparison to other ceratodontids we have studied. Prearticular tooth plates also bear denticles on the crests, but they are less distinct and numerous (e.g., two are present on C2 of BYU 20813, Fig. 3.4) than on pterygopalatine plates. Four major crests are present, but the last is invariably twinned into two well-defined crests, giving a total of five. On the inferior side of the prearticular (Fig. 3.5), Ruge’s Ridge is developed underneath and medial to the C1–C2 ridges, as is commonly seen among North American ceratodontids.
Etymology
Named in honor of Dr. James Ian Kirkland, in recognition of his substantial contributions to the study of Mesozoic lungfishes and to knowledge of the Cedar Mountain Formation and its contained fossils.
Materials
Seven tooth plates, including the holotype, as follows: BYU 20807, an incomplete right prearticular plate with partial prearticular; BYU 20808 (holotype), a complete right pterygopalatine plate with preserved pterygopalatine; BYU 20809, complete left prearticular plate with partial prearticular; BYU 20810, a complete right pterygopalatine plate with partial pterygopalatine; BYU 20811, a complete left pterygopalatine plate with partial pterygopalatine; BYU 20812, an incomplete left prearticular plate, probably from an immature individual; and BYU 20813, a complete left prearticular plate with complete prearticular.
Remarks
Ceratodus kirklandi is only the second North American, Early Cretaceous ceratodontid that is represented by multiple upper and lower plates from the same site. These specimens are probably the basis for listing of “Ceratodus undescribed new species” in the Yellow Cat Member, Cedar Mountain Formation, by Kirkland et al. (Reference Kirkland, Cifelli, Britt, Burge, Decourten, Eaton and Parrish1999, p. 206). Like C. stewarti and Potamoceratodus guentheri, C. kirklandi possesses tall ridges with crest angles (C1Cp) being slightly acute. Ceratodus kirklandi is most similar to C. stewarti from the Moenave and Kayenta formations. Both species have prearticular tooth plates with a twinned fourth ridge crest, a condition unknown in any other North American species of Ceratodus (Milner and Kirkland, Reference Milner and Kirkland2006). Similarly, the pterygopalatine fifth ridge is twinned in C. stewarti (based on the newly referred specimen, OMNH 69332) and variably twinned, at least, in C. kirklandi. Kemp (Reference Kemp1997) argued against the use of these divided ridges as taxonomic characters, based on observations of variable twinning of the last ridge crest in the living Australian lungfish (Neoceratodus forsteri). Our survey of post-Triassic ceratodontids of North America suggests that this condition is often absent, or at least extremely rare, in most species. Here, we follow Milner and Kirkland (Reference Milner and Kirkland2006) and attribute taxonomic significance to this feature.
Ceratodus molossus new species
Holotype
NCSM 30252, a robust, complete right pterygopalatine plate with a partial pterygopalatine attached; Late Cretaceous, Mussentuchit Member, Cedar Mountain Formation; 31 km south of Emery, Emery County, Utah, U.S.A.
Diagnosis
A ceratodontid bearing four primary ridge crests on the pterygopalatine plate, with the last being twinned slightly into an exposed ‘chin’ on the posterolabial corner; mesial margin strongly linear. Both pterygopalatine and prearticular plates have a highly obtuse inner angle (ABC exceeding 130°) in combination with a strongly acute C1Cp angle (69–76°) and nearly planar occlusal surfaces, with negligible development of crests. The prearticular plate is characterized by rounded lingual and posterior margins, and a thick but short C1 ridge crest.
Occurrence
Upper Cretaceous (Cenomanian), Cedar Mountain and Naturita formations, Utah, U.S.A. The referred prearticular specimen (MNA.V.10649) is from MNA locality 1067, 2.4 km northwest of Cannonville, Garfield County, Utah, U.S.A. The referred pterygopalatine plate (UMNH 16774) is also from the Naturita Formation, near Escalante, Garfield County, Utah, U.S.A.
Description
The holotype specimen (NCSM 30252) is a large, right pterygopalatine plate with a complete occlusal surface. An additional specimen, very similar to the holotype, is also described here. UMNH 16774 is a large, right pterygopalatine plate, complete except for the tip of C1. Both known pterygopalatine plates (Fig. 2.2–2.5) strongly resemble the pterygopalatine plate of Ceratodus robustus (Knight, Reference Knight1898, fig. 1), particularly in its large size, extreme flattening of the occlusal surface, shallow and rounded notches between ridges (greatest separation is between C1-C2), and squared posterior margin (Kirkland, Reference Kirkland1987, pl. 1N; 1998, fig. 3I). These plates differs from C. robustus in having a twinned Cp ridge crest; also, ridge crests are thinner, with C3 and Cp ridge crests slightly more distinct (the latter noticeably shorter than in C. robustus). In addition, the elongate, finger-like C1 ridge crest of the holotype specimen is not observed in C. robustus. The large projection seen on the mesiolingual border (junction of inner angle) of C. robustus is also present in NCSM 30252 and UMNH 16774, but is broader and does not project as far lingually.
The prearticular plate referred to Ceratodus molossus (MNA.V.10649, Fig. 2.6, 2.7) is complete; one complete and several partial prearticular plates of C. robustus are available for comparison (Kirkland, Reference Kirkland1987, plate 1P–T; 1998, fig. 3H, J). MNA.V.10649 is slightly smaller than would be expected for the individual represented by the holotype, NCSM 30252, but it is morphologically appropriate and would occlude well with the holotype, were it not for minor deformation of the prearticular tooth plate surface (see below). As with the upper plates, the prearticular tooth plate of C. molossus is similar to that of C. robustus, particularly in its massive construction, negligible expression of ridge crests on the occlusal surface, and the shallowness of the notches between ridges. MNA.V.10649 bears four ridges, of which C1 and C2 are widely spaced; the distance between C2-C3 is slightly greater than between C3-Cp, whereas C2-C4 tend to be more closely spaced, with more angulate intervening notches, in C. robustus. In both, the mesial margin differs from other North American species of Ceratodus in being rounded. In C. molossus, this rounded mesial margin is continuous with similarly curved lingual and posterior margins, giving the plate a semilunate shape. The lingual and posterior margins are straighter in C. robustus. TATE 3000 (Kirkland, Reference Kirkland1998, fig. 3J), the only complete prearticular plate known for C. robustus, is proportionately more labiolingually expanded (especially on the mesial end, where C1 is particularly broad) than MNA.V.10649, but other specimens of C. robustus suggest variability in proportions of the prearticular plate.
Intriguingly, the occlusal surface of MNA.V.10649 is somewhat bowed, with the central portion being concave; a distinct trough (~4 mm wide and 2–3 mm deep) runs labiolingually across the center. These are readily identified as pathologies, of which a number are known for fossil and living lungfishes; the condition in MNA.V.10649 is closely matched in a specimen of Triassic Ceratodus kaupi (Kemp, Reference Kemp2001, fig. 6B; see also Kemp, Reference Kemp2003).
Etymology
From molossus (Latin form of Greek Μολοσσfo), after the extinct breed of Greco-Roman fighting dog (De Prisco and Johnson, Reference De Prisco and Johnson1990), named in reference to Bulldog Bench near Cannonville, Utah, U.S.A., where the prearticular specimen (and other lungfish material) was found. The name is also an allusion to the presumed large and powerful feeding apparatus of the species, reminiscent of that of a bulldog or mastiff.
Materials
Three nearly complete tooth plates, including the holotype: NCSM 30252 (holotype), a right pterygopalatine plate; UMNH 16774, a right pterygopalatine plate; and MNA.V.10649, a right prearticular plate.
Remarks
With the addition of C. molossus, the already diverse ichthyofauna of the Naturita Formation (see Brinkman et al., Reference Brinkman, Newbrey, Neuman and Eaton2013) now includes two species of Ceratodus. Ceratodus gustasoni Kirkland, Reference Kirkland1987, like C. molossus, has flat, low-crested tooth plates; though represented by a number of specimens, only the lower (prearticular) tooth plate is known. It can be readily distinguished from that of C. molossus in that it is proportionately narrower labiolingually, with C1 being elongate and narrow, the crushing surface at the origin of ridge crests more mound-like, the lingual margin straight, and the C2-Cp ridge crests sharply pointed. Ceratodus molossus provides a striking point of contrast with the other recognized Cenomanian species, C. carteri Main, Parris, Grandstaff, and Carter, Reference Main, Parris, Grandstaff and Carter2014, from the Woodbine Formation of Texas, which has small, sharply crested tooth plates.
Ceratodus nirumbee new species
Holotype
OMNH 34765, a complete right prearticular plate, Early Cretaceous (Albian, see Cifelli and Davis, Reference Cifelli and Davis2015), lower unit VII (stratigraphic nomenclature of Ostrom, Reference Ostrom1970), Cloverly Formation; OMNH locality V1061, 11 km east northeast of Edgar, Carbon County, Montana, U.S.A.
Diagnosis
A tall-crested, medium-sized ceratodontid most closely resembling C. fossanovum, differing in the extreme mesial placement of Ruge’s Ridge; ridge crests more distinct, particularly near area of common origin, with narrower crest Cp and better developed C3-Cp notch; and a more angulate, less rounded posterolingual margin.
Occurrence
Known only from the type locality, Cloverly Formation (Albian, Early Cretaceous), Carbon County, Montana, U.S.A.
Description
OMNH 34765 is a complete right prearticular tooth plate with fused prearticular; it is slightly water-worn, lacking the anterior and posterior processes of the prearticular, and has well-developed occlusal wear. The plate bears four tall, well-defined ridge crests and a small crushing surface at the origin of crests C1-C2. Crest C1 bears three denticles in a single row; four denticles in a non-linear arrangement are present on C2; C3-C4 begin with double rows of denticles and have an irregular arrangement. OMNH 34765 represents a medium-sized lungfish, larger than Potamoceratodus guentheri and smaller than Ceratodus gustasoni, most resembling C. fossanovum from the Morrison Formation. OMNH 34765 possesses similar ridge crest angles (Table 1) to C. fossanovum, and falls within the range of variation for this species (see values for C. “guentheri” in Kirkland, Reference Kirkland1987). However, OMNH 34765 differs from C. fossanovum based on suite of characters not found on any of the published specimens from the Jurassic. Ceratodus fossanovum possesses a well-developed crushing platform on the lingual side of prearticular tooth plates, joining the origins of the ridges for the C3 and Cp ridge (Kirkland, Reference Kirkland1987, plate 1D), which is not developed in OMNH 34675. This platform can be seen in lingual view as a second convexity, more posterior to the tall platform formed from the intersection of the C1 and C2 ridges in C. fossanovum. The notch (sulcus) between the C3 and Cp ridge crests is deeper and better developed in OMNH 34765 than in C. fossanovum. On the inferior surface of the underlying prearticular (Fig. 2.9), OMNH 34765 has a well-developed median trough or groove that terminates mesially at Ruge’s Ridge, which is more distally placed in other species.
Etymology
From the Crow, Nirumbee, Little People of the Pryor Mountains; dwarves or devils with a ferocious temperament, sharp teeth, and a predilection for meat (Murray, Reference Murray2008). Allusion is to the presumed feeding habits of the species and to proximity of the type locality to the Pryor Mountains, specifically what is now Chief Plenty Coups State Park, where the legendary Crow leader drew inspiration from the Nirumbee (Linderman, Reference Linderman1957).
Remarks
Ceratodus nirumbee cannot be readily confused with the other ceratodontid known from the Cloverly Formation, C. frazieri Ostrom, Reference Ostrom1970, which is characterized by large, flat-crowned tooth plates (Fig. 2.12–2.13). Oreska et al. (Reference Oreska, Carrano and Dzikiewicz2013) mentioned two ceratodontids among the specimens they collected via underwater screen washing of rock matrix from the Cloverly Formation: C. frazieri and Ceratodus cf. P. guentheri. The specimens are too fragmentary to be identified with confidence, but USNM 546679 (Oreska et al., Reference Oreska, Carrano and Dzikiewicz2013, fig. 5B) much resembles OMNH 34765, holotype of C. nirumbee, particularly in the presence of a long, thin C3 ridge crest that is subequal to the C2 ridge crest. We provisionally consider this specimen, at least, as belonging to the species. Similarly, Bonde (Reference Bonde2008) and Bonde et al. (Reference Bonde, Varricchio, Jackson, Loope and Shirk2008) briefly described and figured a pterygopalatine tooth plate (VOF-2005-03-2A-001) from the Albian-aged Willow Tank Formation of southern Nevada, similar in shape to the holotype of C. nirumbee. If this specimen does in fact represent an upper plate for C. nirumbee, it vastly increases our knowledge of the morphology for this species. Until more robust comparative studies can be done, we informally attribute this specimen to the species as well.
Ceratodus kempae new species
Holotype
BYU 20814, right pterygopalatine tooth plate and attached pterygopalatine, complete except for C1; Early Cretaceous (Valanginian, see Hendrix et al., Reference Hendrix, Moeller, Ludvigson, Joeckel and Kirkland2015); BYU locality 1783, 16.7 km southwest of Cisco, Grand County, Utah, U.S.A.
Diagnosis
Moderate-sized species of Ceratodus, slightly smaller than C. gustasoni, which it resembles in having little surface relief on tooth plate crowns, lack of denticles on ridge crests, obvious circumdenteonal islands present on occlusal surface, and ridges that narrow and are pointed labially. Differs from C. gustasoni and other flat-plated North American species of Ceratodus in having less obtuse C1-Cp angle, slightly better-developed ridge crests, serrated labial plate margin (angulate ridge termini separated by angulate notches), and presence of prominent vertical wear facets between ridges.
Occurrence
Known only from the type locality, Yellow Cat Member, Cedar Mountain Formation (Valanginian, Lower Cretaceous), Grand County, Utah, U.S.A.
Description
BYU 20814 is missing C1 of what was originally a five-crested right pterygopalatine tooth plate. The preserved part of the mesial margin indicates angle ABC to be 111° (Table 1), less than the flat-crowned C. gustasoni or C. frazieri (Kirkland, Reference Kirkland1987, table 1). The specimen is of moderate size, considerably smaller than in C. frazieri and larger than P. guentheri or C. kirklandi. The occlusal surface is low and undulating, with a modestly developed central mound, where the ridges originate at the mesiolingual corner of the plate. Ridge crests are faint but are better developed than in the most closely similar taxon, C. gustasoni (Fig. 2.14, 2.15). All of the labial ridge tips are distinctly angular, as are the intervening notches, such that the labial margin of the plate has a serrated appearance—an appearance enhanced by the fact that the tips lie in a linear configuration (rather than rounded, as is typically the case). Also, the Cp tip is distinctive in being elongate and slender. The occlusal surface is worn and much resembles that of other flat-crowned plates, such as those of C. frazieri or C. gustasoni, in having a cratered appearance owing to the presence of circumdenteonal dentin islands. Wear is also distinct on the near-vertical surfaces in the notches between ridges. The dorsal surface of the fossil reveals the presence of a now-broken but originally large pterygopalatine process above C2-C3, similar to the condition in OMNH 04033, referred to C. frazieri (Kirkland, Reference Kirkland1998), and differing from C. kranzi, in which it lies above C3-C4 (Frederickson et al., Reference Frederickson, Lipka and Cifelli2016).
Etymology
For Anne Kemp, in recognition of her many contributions to understanding the evolution and biology of lungfishes, past and present.
Remarks
Ceratodus kempae is clearly a flat-crested lungfish, with tooth plates that presumably emphasized crushing function (Parris et al., Reference Parris, Grandstaff and Banks2014): the occlusal aspect of the tooth plate has little topography, ridges lack denticles, and the surface has a pock-marked texture owing to dentin structure. On the other hand, C. kempae resembles sharp-crested species such as P. guentheri and C. kirklandi in its relatively small size, low inner (ABC) angle, presence of ridge crests (though faint), and slender, angulated labial ridge tips that are separated by angular notches. In these respects, C. kempae is structurally intermediate between flat-crowned and sharp-crested (“ceratodont” and “ptychoceratodont,” Parris et al., Reference Parris, Grandstaff and Banks2014, p. 279) morphologies. This combination of characters is novel in the North American record, prompting us to give it formal taxonomic recognition.
Three other fragments of lungfish tooth plates have thus far been recovered from the surface at BYU locality 1783. These are small, suggesting that they may belong to juvenile individuals, and are too fragmentary for confident identification, but the presence of sharp ridges on each suggests that they may belong to C. kirklandi, also known from the Yellow Cat Member of the Cedar Mountain Formation.
Discussion
North American ceratodontids in space and time
Given the general scarcity of dipnoan fossils in the Jurassic–Cretaceous of North America, coupled with intraspecific variability and low information content of most specimens (tooth plates), some identifications and taxonomic issues are problematic. Formal systematic revision cannot be attempted here (nor do we believe that revision will be worthwhile until substantial new discoveries are made). In its stead, we provide a brief review of fossil occurrences, as a basis for discussion of diversity patterns through time. The review is arranged by geologic age; lacking a repeatable systematic framework, species are listed alphabetically within each epoch (Table 2). One occurrence is noted in this introductory paragraph because age and provenance are uncertain. This is UNSM 50993, a gigantic (maximum dimension=117 mm) pterygopalatine plate described by Shimada and Kirkland (Reference Shimada and Kirkland2011). The specimen, which undoubtedly represents a distinct, unnamed species of flat-crowned ceratodontid, was found in the outcrop belt of Miocene- to Pliocene-age deposits of the Ogallala Formation in western Nebraska, and was probably transported from a Jurassic or Cretaceous unit to the west.
Table 2 Post-Triassic North American lungfish recognized in this study; C.=Ceratodus; P.=Potamoceratodus.

Early Jurassic
North America’s earliest ceratodontids are of Early Jurassic age; two taxa are currently known, one not positively identified. Ceratodus stewarti Milner and Kirkland, Reference Milner and Kirkland2006 was originally recognized on the basis of a single prearticular plate (UMNH 16027) from the Whitmore Point Member of the Moenave Formation, Utah. The only known pterygopalatine plate, described herein, hails from the stratigraphically higher Kayenta Formation, also in Utah. The species generally resembles Potamoceratodus guentheri (below), although it appears to be more closely related to Early Cretaceous C. kirklandi n. sp. The known range of C. stewarti is Hettangian–Sinemurian.
The other Early Jurassic occurrence is an unidentified taxon, Potamoceratodus cf. P. guentheri, based on MCZ 13865, a prearticular plate distinguished from the otherwise similar Ceratodus stewarti in having higher crests and more acute angulation of C1Cp (Milner and Kirkland, Reference Milner and Kirkland2006), as well as lacking the twinned Cp crest on the lower plates. The specimen is from the Kayenta Formation of northern Arizona.
Late Jurassic
No ceratodontids are yet recorded from the Middle Jurassic, but the Morrison Formation (Kimmeridgian–Tithonian) has yielded four species. Ceratodus americanus Knight, Reference Knight1898 is represented by UWM 2001b, a right pterygopalatine plate lacking C1. Kirkland (Reference Kirkland1987) considered C. americanus a junior subjective synonym of Potamoceratodus guentheri (Marsh, Reference Marsh1878), based on specimens incorrectly referred to the latter species. Correcting this error, he later (Kirkland, Reference Kirkland1998) transferred those specimens to a new species, Ceratodus fossanovum Kirkland, Reference Kirkland1998 (misspelling in the original; intended naming for Como Quarry 9 would be fossanovem). Given the earlier referral of UWM 2001b, it is logical to wonder whether C. americanus might be a senior subjective synonym of C. fossanovum, which it closely resembles. Considering the incompleteness of UWM 2001b, we defer this taxonomic pitfall, and provisionally regard C. americanus as a nomen dubium (see Mones, Reference Mones1989) for present purposes.
Ceratodus fossanovum is the best-represented (in terms of number of specimens) and most stratigraphically restricted of Ceratodontidae from the Morrison Formation: it is unequivocally known only from Quarry 9 (see Simpson, Reference Simpson1926; Carrano and Velez-Juarbe, Reference Carrano and Velez-Juarbe2006) and nearby, stratigraphically equivalent sites in the Como Bluff area, Wyoming. Available fossils show size variation, perhaps related to ontogenetic age. Ceratodus fossanovum resembles P. guentheri, differing in its larger size, more obtuse inner angle (ABC), and presence of a crushing platform on tooth plates. In these respects, it is morphologically closest to Early Cretaceous C. nirumbee n. sp.
The record of Ceratodus frazieri Ostrom, Reference Ostrom1970 in the Morrison Formation is based on a lower tooth plate and nearly complete prearticular (SDSM 426) described by Pinsof (Reference Pinsof1983). As noted by Kirkland (Reference Kirkland1987), it is strikingly similar to specimens from the Cloverly Formation (Albian, Early Cretaceous; see Fig. 2.12, 2.13, and Supplemental Data 8), including the holotype, YPM 5276. OMNH 04033, a pterygopalatine plate from the upper Morrison Formation of western Oklahoma, was also provisionally identified as C. frazieri (see Kirkland, Reference Kirkland1998, fig. 3G). The specimen is broadly similar to KUVP 16226, an upper tooth plate from the Kiowa Shale (Lower Cretaceous) of Kansas, referred to C. frazieri by Schultze (Reference Schultze1981). Both are rather flat, with a slightly excavated (concave) occlusal surface, and have relatively wide notches between the ridges. OMNH 04033 is somewhat smaller and is labiolingually narrower, with a more tapering, less quadrate posterior margin, and with a somewhat thicker crown and distinct labial ridge crests. Whether these differences are ontogenetic or taxonomic cannot be determined with material in hand; we accept Kirkland’s (Reference Kirkland1998) referral for purpose of this review.
Potamoceratodus guentheri (Marsh, Reference Marsh1878) (same as, or including Ceratodus felchi Kirkland, Reference Kirkland1987) was the first dipnoan to be named from the Morrison Formation. It is a high-crested form with slightly acute C1Cp angle, and is among the smallest of North America’s post-Triassic lungfish; it is also notable in being the only North American ceratodontid known by cranial material (Pardo et al., Reference Pardo, Huttenlocker, Small and Gorman2010). A reasonable sample of tooth plates is known from Felch Quarry in the lower part of the Morrison Formation (Kirkland, Reference Kirkland1987). Potamoceratodus guentheri is also known from geographically and stratigraphically scattered sites in the unit (Kirkland, Reference Kirkland1998), and it, or a closely similar species, is represented in the Early Jurassic of Arizona. Two species with morphologically similar tooth plates are also known from the Cretaceous of Texas (Main et al., Reference Main, Parris, Grandstaff and Carter2014; Parris et al., Reference Parris, Grandstaff and Banks2014).
Ceratodus robustus Knight, Reference Knight1898 is represented by a small sample that nonetheless includes all upper (pterygopalatine, vomerine) and lower (prearticular) tooth plates, as well as specimens exhibiting a range of variation (Kirkland, Reference Kirkland1987, Reference Kirkland1998). Ceratodus robustus is known from several sites, all in the Morrison Formation of Wyoming. Until now, it has been regarded as unique among North American species of Ceratodus, and has been occasionally cited in connection with possible relatives from other landmasses (e.g., Kirkland, Reference Kirkland1987, Reference Kirkland1998; Kemp, Reference Kemp1993; Pardo et al., Reference Pardo, Huttenlocker, Small and Gorman2010). Ceratodus molossus n. sp. is closely similar, and suggests that the clade survived into the Late Cretaceous.
Early Cretaceous
We recognize six species of Ceratodus from geographically and stratigraphically scattered sites in the Early Cretaceous of North America. Of these, C. kirklandi n. sp., C. kempae n. sp. (both from the Valanginian of Utah), and C. nirumbee n. sp. (Albian, Montana) are described herein and need no further comment. Ceratodus frazieri Ostrom, Reference Ostrom1970, the first ceratodontid to be named from the Early Cretaceous of North America, was recognized on the basis of a left prearticular plate, YPM 5276 (Ostrom, Reference Ostrom1970, plate 9A), from unit V of the Cloverly Formation, Wyoming. Kirkland (Reference Kirkland1987, plate 1F) illustrated another prearticular plate (MOR 367). Oreska et al. (Reference Oreska, Carrano and Dzikiewicz2013) referred several incomplete tooth plates to C. frazieri. The most complete and the only of these to be illustrated, USNM 546680 (Oreska et al., Reference Oreska, Carrano and Dzikiewicz2013, fig. 5A), is consistent with C. frazieri in being low crowned with rather robust, short ridges, though it is not sufficiently complete for secure identification. Interestingly, it is small (maximum preserved length=26 mm), and may represent an immature individual. An additional specimen (OMNH 60408; Fig. 2.12, 2.13, and Supplemental Data 8) from the Cloverly Formation at OMNH locality V1075 in Big Horn County, Montana, resembles the holotype and can unambiguously be referred to C. frazieri. Schultze (Reference Schultze1981) referred to C. frazieri a large, flat-crowned pterygopalatine plate (KUVP 16262) from the Kiowa Formation (Albian) of western Kansas. The age and morphology of the specimen are appropriate for the species, and we follow Kirkland (Reference Kirkland1987, Reference Kirkland1998) in accepting this referral. Kirkland (Reference Kirkland1987) also suggested possible referral of UCM 49389 (large tooth plate fragments) to this species. The fossils are said to come from the lower part of the Cedar Mountain Formation, central Utah. This unit includes several members, collectively spanning much of the Early Cretaceous and extending to (or just past) the Early-Late Cretaceous boundary (e.g., Kirkland et al., Reference Kirkland, Britt, Burge, Carpenter, Cifelli, Decourten, Eaton, Hasiotis and Lawton1997, Reference Kirkland, Cifelli, Britt, Burge, Decourten, Eaton and Parrish1999). We have not studied the material, and given that the specimens are incomplete and were not illustrated, they can be ignored for present purposes.
Ceratodus kranzi Frederickson, Lipka, and Cifelli, Reference Frederickson, Lipka and Cifelli2016 is based on an incomplete pterygopalatine plate, USNM 508543, from the Arundel Clay facies (Aptian–Albian) of the Potomac Formation, Maryland. This specimen represents a large, robust-plated ceratodontid, most similar in appearance to KUVP 16262 (see Schultze, Reference Schultze1981) from the Kiowa Shale. USNM 508543 is one of only two described Cretaceous dipnoan fossils from eastern North America, and the only one from the Early Cretaceous.
Thurmond (Reference Thurmond1974) reported a sharp-crested species of Ceratodus in the Butler Farm local fauna, Paluxy Formation, Texas. He compared then-available specimens favorably to C. americanus (herein regarded as a nomen dubium) from the Morrison Formation. Kirkland (Reference Kirkland1987) regarded the Trinity species as an unnamed relative of what he later (Kirkland, Reference Kirkland1998) described as C. fossanovum, also from the Morrison Formation. The Trinity species was formally described as Ceratodus texanus Parris, Grandstaff, and Banks, Reference Parris, Grandstaff and Banks2014, based on eight specimens (four pterygopalatine and four prearticular tooth plates) from localities in the Twin Mountains and Paluxy formations (Aptian–Albian, see Winkler et al., Reference Winkler, Murry and Jacobs1990), north-central Texas. Parris et al. (Reference Parris, Grandstaff and Banks2014) compared C. texanus favorably with both Potamoceratodus guentheri and C. fossanovum, distinguishing the species on the basis of size and differences in angles formed by major crests.
Late Cretaceous
Non-marine vertebrates of North America’s Late Cretaceous are incomparably better sampled than those of the Early Cretaceous (e.g., Kielan-Jaworowska et al., Reference Kielan-Jaworowska, Cifelli and Luo2004; Weishampel et al., Reference Weishampel, Dodson and Osmólska2004; Benson et al., Reference Benson, Mannion, Butler, Upchurch, Goswami and Evans2013). In this context, the fact that only four ceratodontids are known for the Late Cretaceous suggests a real decline in diversity of the group. (Two additional taxa from the Late Cretaceous, Ceratodus cruciferus Cope, Reference Cope1876 and C. hieroglyphus Cope, Reference Cope1876, were transferred to Acipenser by Estes [Reference Estes1964]). Three of the four are of Cenomanian (early Late Cretaceous) age. Ceratodus molossus n. sp., a large species with flat-crowned tooth plates (described herein), is from the Cedar Mountain and Naturita formations of Utah and is a probable relative of Late Jurassic C. robustus. Ceratodus gustasoni Kirkland, Reference Kirkland1987, somewhat smaller but also flat-crowned, also comes from the Cenomanian of Utah. The holotype, UCM 54074, and paratype, UCM 54073, are both left prearticular plates. Although other specimens have since been discovered (Fig. 2.14, 2.15, and Supplemental Data 9), pterygopalatine plates for this species are still unknown. Fossils of juveniles have been referred to the species (Brinkman et al., Reference Brinkman, Newbrey, Neuman and Eaton2013, fig. 10.30), but these are uninformative. Kirkland (Reference Kirkland1987) hypothesized that C. gustasoni may be closely related to C. frazieri, differing most noticeably from this species by its posteriorly expanded lingual margin. A specimen of a juvenile, identified as Ceratodus sp. indet. and recently described from a Santonian level in the Iron Springs Formation, may belong to C. gustasoni (Eaton et al., Reference Eaton, Gardner, Kirkland, Brinkman and Nydam2014). Regardless, the specimen is of interest because it and an undescribed species from Los Peyotes, Coahuila Mexico (Coniacian–Santonian) (González-Rodríguez et al., Reference González-Rodríguez, Fielitz, Bravo-Cuevas and Baños-Rodríguez2016) represent the geologically youngest published ceratodontids from the Western Interior. However, putative lungfish remains have also been discovered from the lower Campanian part of the Aguja Formation of Western Texas (Wick et al., Reference Wick, Lehman and Brink2015; personal communication, A.A. Brink, 2016). If confirmed through future study and publication, this material would significantly extend the known range of lungfish from Laramidia (the western part of North America, separated from Appalachia by the Western Interior during much of the Late Cretaceous; Archibald, Reference Archibald1996).
Ceratodus carteri Main, Parris, Grandstaff, and Carter, Reference Main, Parris, Grandstaff and Carter2014 is represented by a relatively large sample, with seven known tooth pates (one pterygopalatine and the remainder prearticulars) from the Woodbine Formation (Cenomanian) of Tarrant County, Texas. The sample also includes a wide size range, presumably reflecting different ontogenetic ages. In general, this species is most similar to the tall-crested P. guentheri. Ceratodus carteri also resembles tall-crested C. texanus (as noted by Parris et al., Reference Parris, Grandstaff and Banks2014), C. nirumbee (both Early Cretaceous), and C. fossanovum (Late Jurassic), but differs in its smaller size, more obtuse C1Cp angle, and lack of a crushing platform.
A right prearticular and overlying plate (NJSM 18774), found at the Big Brook locality in Monmouth County, New Jersey, represents the geologically youngest occurrence of Ceratodontidae in North America, and is one of only two specimens from the eastern seaboard of the continent. Specimens from the Big Brook locality probably come from the Mount Laurel Formation and are of Campanian age (Gallagher et al., Reference Gallagher, Parris and Spamer1986; Lauginiger, Reference Lauginiger1986). NJSM 18774 is structurally indistinguishable from prearticular plates of Ceratodus frazieri from the Cloverly Formation. Parris et al. (Reference Parris, Grandstaff and Gallagher2004) identified the Big Brook specimen as Ceratodus sp. aff. C. frazieri, commenting that it probably represents a different species owing to its occurrence. We do not disagree, but because there is no anatomical basis for this distinction, we simply refer the specimen to C. frazieri without issue. Just as Ceratodus is regarded as a form genus among Ceratodontidae (e.g., Schultze, Reference Schultze1981), so might C. frazieri well be taken as a form species within that genus.
Diversity and paleoecology
As noted, the nature of the fossil record of post-Triassic lungfishes in North America—which consists almost entirely of rather simple tooth plates—places serious constraints on phylogenetic and paleobiologic inference. In this case, the path of reasoned commentary lies perilously close to a chasm of fantasy. These caveats notwithstanding, we find ourselves largely in agreement with phylogenetic and ecomorphologic interpretations proposed by previous authors (Kirkland, Reference Kirkland1987, Reference Kirkland1998; Parris et al., Reference Parris, Grandstaff and Banks2014), although we are unconvinced that existing data support recognition of ancestor-descendant relationships. The welcome new additions to the fossil record help span temporal and geographic gaps, and add considerably to diversity of North American Mesozoic ceratodontids. We propose a simple model, consisting of four species groups, to serve as a framework for discussing the post-Triassic evolutionary history of dipnoans in North America.
As noted by Kirkland (Reference Kirkland1987), tooth plates of North American ceratodontids span a morphological continuum, from small, high-crested Potamoceratodus guentheri to large, flat-crowned Ceratodus robustus. Our four species groups are named for the four species from the Upper Jurassic Morrison Formation: P. guentheri, C. fossanovum, C. frazieri, and C. robustus. The categories are phenetic: we do not intend these groups as definable phylogenetic units, although in most cases a close relationship of within-group species represents a plausible working hypothesis. Broadly speaking, at least, it is also reasonable to suppose that these groups reflect ecomorphological differences among species. Small-bodied taxa with high-crested tooth plates, suggestive of some cutting/slicing ability, are presumed to have relied on soft dietary fare, perhaps including invertebrates; whereas larger ceratodontids having heavily built, flat tooth plates may have had a more varied menu, including shelled prey (Kirkland, Reference Kirkland1987; Bakker, Reference Bakker2009).
Following our grouping scheme, the earliest post-Triassic dipnoans in North America are both members of the P. guentheri group (C. stewarti and Potamoceratodus cf. P. guentheri; Milner and Kirkland, Reference Milner and Kirkland2006; this study) from the Early Jurassic of the southwest (Moenave and Kayenta formations). Intriguingly, tooth plates of C. stewarti bear a twinned Cp ridge, a feature also seen in Early Cretaceous C. kirklandi (Valanginian, Utah), suggesting the possibility of a sub-clade within the P. guentheri group—which itself may or may not be monophyletic. By the Late Jurassic, diversity vastly increases, with all four groups co-occurring in the Morrison Formation. Given the current gap in knowledge for terrestrial ecosystems of the Middle Jurassic of North America (not to mention poor anatomical representation of fossil lungfishes generally), it is currently unknown whether any of these novel groups are derived from the P. guentheri group (Parris et al., Reference Parris, Grandstaff and Banks2014) or immigrants from other continents. Significantly, all of this additional diversity involves taxa characterized by larger size and increased crushing capacity. Most distinctive in this regard is C. robustus, which has been compared favorably with C. kaupi Agassiz, Reference Agassiz1838 (Triassic, Europe) and C. diutinus Kemp, Reference Kemp1993, which ranges from the Aptian–Miocene of Australia (Kemp, Reference Kemp1993). Assuming non-convergent within-group relationships, all four species groups present in the Morrison Formation survived into the Albian, after which the C. fossanovum group disappears, followed by the P. guentheri group and the C. robustus group after the Cenomanian (Fig. 4). Unquestionably the most noteworthy recent improvement in the record of North American ceratodontids is in the medial Cretaceous (Albian–Cenomanian), with the addition of four species, collectively representing three of the four groups (Main et al., Reference Main, Parris, Grandstaff and Carter2014; Parris et al., Reference Parris, Grandstaff and Banks2014; this study). Among these, the most surprising is the Cenomanian C. molossus n. sp., which appears to be a close relative of C. robustus, and hence represents the only record of the C. robustus group outside of the Morrison Formation of Wyoming.

Figure 4 Ceratodus diversity through the Cretaceous of North America, based on the four Jurassic forms (bottom). Major morphotypes (‘Ptychoceratodont’ and ‘Ceratodont,’ top) after Parris et al. (Reference Parris, Grandstaff and Banks2014). Colored columns represent species groups discussed in the text (also indicated by name at base of each column); the form diversity column, right, represents total number of species groups present in given interval (epoch, denoted by dashed horizontal lines). (1) C. kirklandi n. sp.; (2) C. kempae n. sp.; (3) C. kranzi; (4) C. texanus; (5) C. nirumbee n. sp.; (6) C. frazieri; (7) C. frazieri; (8) C. carteri; (9) C. gustasoni; (10) C. molossus n. sp.; (11) C. gustasoni (no images of diagnostic fossil available); (12) C. frazieri. Silhouettes modified from images in Kirkland (Reference Kirkland1987), Parris et al. (Reference Parris, Grandstaff and Gallagher2004, Reference Parris, Grandstaff and Banks2014), Main et al. (Reference Main, Parris, Grandstaff and Carter2014), and Frederickson et al. (Reference Frederickson, Lipka and Cifelli2016). Time scale from Cohen et al. (Reference Cohen, Finney, Gibbard and Fan2013). Specimens from the Willow Tank Formation, Nevada; Los Peyotes, Mexico; and the Aguja Formation, Texas are mentioned in the discussion, but not shown in this figure.
The existing record, including these new occurrences, suggests that lungfish diversity in North America remained relatively stable from the Late Jurassic through the Cenomanian—an interval of ~60 Ma (time scale after Cohen et al., Reference Cohen, Finney, Gibbard and Fan2013) (Fig. 4). To date, geologically younger horizons have yielded only two described specimens: one from the probable Santonian of Utah (Eaton et al., Reference Eaton, Gardner, Kirkland, Brinkman and Nydam2014) and the other from the Campanian of New Jersey (Parris et al., Reference Parris, Grandstaff and Gallagher2004). Both belong to the C. frazieri group; the last-surviving member presumably occurred in a refugium on the east coast of Appalachia, which was then separated from Laramidia (and its well-known Late Cretaceous vertebrate faunas) by the Western Interior Seaway. Given the general scarcity of dipnoan fossils and inadequate knowledge of early Late Cretaceous assemblages, little more can be said about the decline and extinction of lungfishes in North America. It is worthwhile noting, however, that the major drop in abundance and diversity occurs within the context of a broader turnover event among aquatic taxa at the Cenomanian-Turonian boundary (Eaton et al., Reference Eaton, Kirkland, Hutchison, Denton, O’Neill and Parrish1997; Brinkman et al., Reference Brinkman, Newbrey, Neuman and Eaton2013).
North American Ceratodus species are known from a variety of aquatic environments. Most commonly found in freshwater lake or stream deposits (Kirkland, Reference Kirkland1987), it is likely that many of these species had lifestyles similar to the modern Australian lungfish (Neoceratodus forsteri), inhabiting diverse freshwater waterways, but preferring straight, flowing streams and rivers bordered by heavy vegetation and containing woody debris (Kemp, Reference Kemp1986; Arthington, 2009). Although often found in a range of low-quality habitats, members of this species are largely salt intolerant, incapable of living in or migrating through saline waters (Arthington, Reference Arthington2008). Similar to the modern Neoceratodus, Jurassic North American ceratodontids appear to be limited to terrestrial environments. The Early Jurassic species (P. guentheri group) are only known from freshwater deposits in the Moenave and Kayenta formations. The holotype of C. stewarti, for example, was discovered as part of a relatively diverse lacustrine fauna (Milner and Kirkland, Reference Milner and Kirkland2006). Similarly, Late Jurassic species are found in terrigenous rocks deposited under generally arid conditions (Engelmann et al., Reference Engelmann, Chure and Fiorillo2004), with most fossils deriving from fluvial (Kirkland, Reference Kirkland1987) or lacustrine (Pardo et al., Reference Pardo, Huttenlocker, Small and Gorman2010) facies, far from marine habitats (Fig. 5.1).

Figure 5 Maps showing approximate North American lungfish distribution during the: (1) Kimmeridgian to Tithonian, Late Jurassic (Kirkland, Reference Kirkland1987, Reference Kirkland1998; Pardo et al., Reference Pardo, Huttenlocker, Small and Gorman2010); (2) Valanginian to Albian, Early Cretaceous (Ostrom, Reference Ostrom1970; Schultze, Reference Schultze1981; Kirkland, Reference Kirkland1987; Oreska et al., Reference Oreska, Carrano and Dzikiewicz2013; Parris et al., Reference Parris, Grandstaff and Banks2014; Frederickson et al., Reference Frederickson, Lipka and Cifelli2016); (3) Cenomanian, Late Cretaceous (Kirkland, Reference Kirkland1987; Main et al., Reference Main, Parris, Grandstaff and Carter2014); (4) Santonian to Campanian, Late Cretaceous (Parris et al., Reference Parris, Grandstaff and Gallagher2004). Numbers and color grouping correspond to Figure 4; abbreviations for species groups (1): Fr, Ceratodus frazieri; Fo, C. fossanovum; Gu, Potamoceratodus guentheri; and Ro, C. robustus. Maps modified from © Ron Blakey, Colorado Plateau Geosystems, and were chosen based on their best fit to the depositional environments observed at all of the fossil sites, combined with the closest temporal representation available. Specimens from the Willow Tank Formation, Nevada; Los Peyotes, Mexico; and the Aguja Formation, Texas are mentioned in the discussion but not shown in this figure. Scale bar=200 km.
By contrast, Early Cretaceous occurrences (Fig. 5.2) of North American Ceratodontidae occur in a wider range of depositional settings, varying from fluvial (Ostrom, Reference Ostrom1970) to nearshore marine (Schultze, Reference Schultze1981). Species from the Late Cretaceous (Fig. 5.3, 5.4) all occur in close proximity to the paleo-shoreline, and several sites lie in horizons that were probably deposited under brackish or marine settings. Within the C. frazieri group, for example, at least two specimens derived from nearshore marine deposits (Schultze, Reference Schultze1981; Parris et al., Reference Parris, Grandstaff and Gallagher2004), and one is from a possibly paludal setting (Frederickson et al., Reference Frederickson, Lipka and Cifelli2016). Other large ceratodontids may have also had some degree of salt tolerance. For example, fossils of C. molossus (C. robustus group) and C. gustasoni (C. frazieri group) were found in the near-coastal sediments of the Naturita Formation (Kirkland, Reference Kirkland1987; this study). Similarly, a Late Cretaceous member of the P. guentheri group (C. carteri) is known from a deltaic plain or coastal wetland environment (Main et al., Reference Main, Parris, Grandstaff and Carter2014), suggesting that salt tolerance may have developed among gracile species groups as well. Notably, the P. guentheri and C. frazieri groups are also the only dipnoans known from Appalachia during the Cretaceous, although it is likely that they inhabited the area prior to completion of the Western Interior Seaway. At least one Australian species of Ceratodus, wide-ranging C. diutinus, occurs in both freshwater and marine units (Kemp, Reference Kemp1993). Ceratodus also occurs in the marine Westbury Formation (Late Triassic) of Britain, but the vertebrate fossils from this unit apparently represent a mixed assemblage (Storrs, Reference Storrs1993), so the origin of the lungfish specimens is unclear.
Alternatively, it could be argued that all of these later brackish to saltwater occurrences represent reworked material from freshwater settings. Indeed, the Campanian C. frazieri fossil from New Jersey was said to be “taphonomically mature” (Parris et al., Reference Parris, Grandstaff and Gallagher2004, p. 65), indicating that this specimen was transported prior to final burial. In most cases, however, reworking hypotheses are based on behavioral observations of salt intolerant modern lungfish and not paleontological evidence (Kirkland, Reference Kirkland1987; Kemp, Reference Kemp1993). In isolated cases, the reworking hypothesis may seem more parsimonious, but taken as a whole, the Ceratodus record of the Late Cretaceous of North America appears to include more coastal, or at least coastal adjacent, settings, than that of the preceding Jurassic.
Besides a functional lung, Dipnoi possess a variety of traits that allow them to survive in an array of environments, including ephemeral bodies of water. For example, aestivation allows living lepidosirenids (Lepidosiren and Protopterus) to lie dormant in an underground cocoon of mucus during drought. Trace fossils interpreted as aestivation chambers belonging to the basal dipnoan Gnathorhiza can be locally quite common in Paleozoic rocks of North America (e.g., Carlson, Reference Carlson1968; Berman, Reference Berman1976), implying that this behavior has a deep history within the evolution of the clade. The most basal living species of lungfish (Neoceratodus forsteri), however, does not aestivate. By analogy, and noting the absence of identified aestivation burrows in geologically younger rocks, Kirkland (Reference Kirkland1987; see also Bakker and Bir, Reference Bakker and Bir2004) suggested that post-Triassic North American lungfish did not aestivate either. Conversely, in their phylogeny of Dipnoi, Cavin et al. (Reference Cavin, Suteethorn, Buffetaut and Tong2007) hypothesized that all taxa more derived than Neoceratodus (including Ceratodus) would have been exclusively freshwater fish that were capable of aestivating during dry spells. Burrows of the correct size and shape to accommodate a full-grown Ceratodus would hypothetically be conspicuous, especially for some of the larger species, but to date none has been found. Thus we tentatively reject the hypothesis of Cavin et al. (Reference Cavin, Suteethorn, Buffetaut and Tong2007) because: (1) North American ceratodontids may not have been limited to freshwater habitats, and (2) there is no evidence for dipnoan aestivation chambers in the Jurassic and Cretaceous of North America. The second point also argues against an alternative hypothesis: that marine-dwelling Ceratodus may have burrowed during low tide (see Schultze and Chorn, Reference Schultze and Chorn1997).
Conclusions
In the three decades since the last systematic treatment of North American ceratodontids (Kirkland, Reference Kirkland1987), the number of recognized species has more than doubled, from five to 12, with most of the new taxa coming from the middle part of the Cretaceous (Albian–Cenomanian). Still, few of these species are known from more than a few specimens, and only one is represented by material other than tooth plates and the bones that bear them. This limitation still makes direct phylogenetic study difficult, and it remains debatable as to whether included species belong to single or multiple genera (see Pardo et al., Reference Pardo, Huttenlocker, Small and Gorman2010). Nonetheless, a phenetic approach helps organize species into useful species groups, which, in some cases at least, likely represent clades. Using this technique, it is apparent that all of the Late Jurassic species have at least one morphological counterpart in the Early Cretaceous. Similarly, present evidence points to a significant extinction event during the Cenomanian–Turonian, exterminating all but the C. frazieri group. More work is needed to pinpoint the cause of this extinction, as well as to determine the relationships among post-Triassic North American lungfishes. Here, as in so many other cases, the most urgent need is for basic data—a need that can only be addressed by further field work.
Acknowledgments
For access to specimens in their care, we thank C. Beard, D. Burnham, M. Florence, J. Whitmore Gillette, L. Herzog, J. Krishna, R. Scheetz, V. Schneider, S. Underwood, and L. Zanno. We are grateful to A. Titus and D. Parris for various bits of information, and to R. Burkhalter and J. Cohen for help with photography and 3D digitalization. We also thank B. Grandstaff and L. Cavin for their thoughtful reviews, which greatly benefited this work. Partial support for this research was provided by grants from the National Geographic Society (4761-91, 5021-92, 5918-97, and W266-13) and the National Science Foundation (BSR 8906992, DEB 9401994, 9870173).
Accessibility of supplemental data
As supplemental data, we provide 3D PDF files for most of the specimens illustrated herein. Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.v1jv1







