We introduce a modification of the generalized Pólya urn model containing two urns, and we study the number of balls  $B_j(n)$ of a given color
$B_j(n)$ of a given color  $j\in\{1,\ldots,J\}$ added to the urns after n draws, where
$j\in\{1,\ldots,J\}$ added to the urns after n draws, where  $J\in\mathbb{N}$. We provide sufficient conditions under which the random variables
$J\in\mathbb{N}$. We provide sufficient conditions under which the random variables  $(B_j(n))_{n\in\mathbb{N}}$, properly normalized and centered, converge weakly to a limiting random variable. The result reveals a similar trichotomy as in the classical case with one urn, one of the main differences being that in the scaling we encounter 1-periodic continuous functions. Another difference in our results compared to the classical urn models is that the phase transition of the second-order behavior occurs at
$(B_j(n))_{n\in\mathbb{N}}$, properly normalized and centered, converge weakly to a limiting random variable. The result reveals a similar trichotomy as in the classical case with one urn, one of the main differences being that in the scaling we encounter 1-periodic continuous functions. Another difference in our results compared to the classical urn models is that the phase transition of the second-order behavior occurs at  $\sqrt{\rho}$ and not at
$\sqrt{\rho}$ and not at  $\rho/2$, where
$\rho/2$, where  $\rho$ is the dominant eigenvalue of the mean replacement matrix.
$\rho$ is the dominant eigenvalue of the mean replacement matrix.
We consider linear-fractional branching processes (one-type and two-type) with immigration in varying environments. For  $n\ge0$, let
$n\ge0$, let  $Z_n$ count the number of individuals of the nth generation, which excludes the immigrant who enters the system at time n. We call n a regeneration time if
$Z_n$ count the number of individuals of the nth generation, which excludes the immigrant who enters the system at time n. We call n a regeneration time if  $Z_n=0$. For both the one-type and two-type cases, we give criteria for the finiteness or infiniteness of the number of regeneration times. We then construct some concrete examples to exhibit the strange phenomena caused by the so-called varying environments. For example, it may happen that the process is extinct, but there are only finitely many regeneration times. We also study the asymptotics of the number of regeneration times of the model in the example.
$Z_n=0$. For both the one-type and two-type cases, we give criteria for the finiteness or infiniteness of the number of regeneration times. We then construct some concrete examples to exhibit the strange phenomena caused by the so-called varying environments. For example, it may happen that the process is extinct, but there are only finitely many regeneration times. We also study the asymptotics of the number of regeneration times of the model in the example.
We consider continuous-state branching processes (CB processes) which become extinct almost surely. First, we tackle the problem of describing the stationary measures on  $(0,+\infty)$ for such CB processes. We give a representation of the stationary measure in terms of scale functions of related Lévy processes. Then we prove that the stationary measure can be obtained from the vague limit of the potential measure, and, in the critical case, can also be obtained from the vague limit of a normalized transition probability. Next, we prove some limit theorems for the CB process conditioned on extinction in a near future and on extinction at a fixed time. We obtain non-degenerate limit distributions which are of the size-biased type of the stationary measure in the critical case and of the Yaglom distribution in the subcritical case. Finally we explore some further properties of the limit distributions.
$(0,+\infty)$ for such CB processes. We give a representation of the stationary measure in terms of scale functions of related Lévy processes. Then we prove that the stationary measure can be obtained from the vague limit of the potential measure, and, in the critical case, can also be obtained from the vague limit of a normalized transition probability. Next, we prove some limit theorems for the CB process conditioned on extinction in a near future and on extinction at a fixed time. We obtain non-degenerate limit distributions which are of the size-biased type of the stationary measure in the critical case and of the Yaglom distribution in the subcritical case. Finally we explore some further properties of the limit distributions.
We investigate some aspects of the problem of the estimation of birth distributions (BDs) in multi-type Galton–Watson trees (MGWs) with unobserved types. More precisely, we consider two-type MGWs called spinal-structured trees. This kind of tree is characterized by a spine of special individuals whose BD  $\nu$ is different from the other individuals in the tree (called normal, and whose BD is denoted by
$\nu$ is different from the other individuals in the tree (called normal, and whose BD is denoted by  $\mu$). In this work, we show that even in such a very structured two-type population, our ability to distinguish the two types and estimate
$\mu$). In this work, we show that even in such a very structured two-type population, our ability to distinguish the two types and estimate  $\mu$ and
$\mu$ and  $\nu$ is constrained by a trade-off between the growth-rate of the population and the similarity of
$\nu$ is constrained by a trade-off between the growth-rate of the population and the similarity of  $\mu$ and
$\mu$ and  $\nu$. Indeed, if the growth-rate is too large, large deviation events are likely to be observed in the sampling of the normal individuals, preventing us from distinguishing them from special ones. Roughly speaking, our approach succeeds if
$\nu$. Indeed, if the growth-rate is too large, large deviation events are likely to be observed in the sampling of the normal individuals, preventing us from distinguishing them from special ones. Roughly speaking, our approach succeeds if  $r\lt \mathfrak{D}(\mu,\nu)$, where r is the exponential growth-rate of the population and
$r\lt \mathfrak{D}(\mu,\nu)$, where r is the exponential growth-rate of the population and  $\mathfrak{D}$ is a divergence measuring the dissimilarity between
$\mathfrak{D}$ is a divergence measuring the dissimilarity between  $\mu$ and
$\mu$ and  $\nu$.
$\nu$.
We investigate branching processes in varying environment, for which  $\overline{f}_n \to 1$ and
$\overline{f}_n \to 1$ and  $\sum_{n=1}^\infty (1-\overline{f}_n)_+ = \infty$,
$\sum_{n=1}^\infty (1-\overline{f}_n)_+ = \infty$,  $\sum_{n=1}^\infty (\overline{f}_n - 1)_+ < \infty$, where
$\sum_{n=1}^\infty (\overline{f}_n - 1)_+ < \infty$, where  $\overline{f}_n$ stands for the offspring mean in generation n. Since subcritical regimes dominate, such processes die out almost surely, therefore to obtain a nontrivial limit we consider two scenarios: conditioning on nonextinction, and adding immigration. In both cases we show that the process converges in distribution without normalization to a nondegenerate compound-Poisson limit law. The proofs rely on the shape function technique, worked out by Kersting (2020).
$\overline{f}_n$ stands for the offspring mean in generation n. Since subcritical regimes dominate, such processes die out almost surely, therefore to obtain a nontrivial limit we consider two scenarios: conditioning on nonextinction, and adding immigration. In both cases we show that the process converges in distribution without normalization to a nondegenerate compound-Poisson limit law. The proofs rely on the shape function technique, worked out by Kersting (2020).
We consider a discrete-time population growth system called the Bienaymé–Galton–Watson stochastic branching system. We deal with a noncritical case, in which the per capita offspring mean 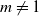 $m\neq1$. The famous Kolmogorov theorem asserts that the expectation of the population size in the subcritical case
$m\neq1$. The famous Kolmogorov theorem asserts that the expectation of the population size in the subcritical case 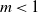 $m<1$ on positive trajectories of the system asymptotically stabilizes and approaches
$m<1$ on positive trajectories of the system asymptotically stabilizes and approaches  ${1}/\mathcal{K}$, where
${1}/\mathcal{K}$, where  $\mathcal{K}$ is called the Kolmogorov constant. The paper is devoted to the search for an explicit expression of this constant depending on the structural parameters of the system. Our argumentation is essentially based on the basic lemma describing the asymptotic expansion of the probability-generating function of the number of individuals. We state this lemma for the noncritical case. Subsequently, we find an extended analogue of the Kolmogorov constant in the noncritical case. An important role in our discussion is also played by the asymptotic properties of transition probabilities of the Q-process and their convergence to invariant measures. Obtaining the explicit form of the extended Kolmogorov constant, we refine several limit theorems of the theory of noncritical branching systems, showing explicit leading terms in the asymptotic expansions.
$\mathcal{K}$ is called the Kolmogorov constant. The paper is devoted to the search for an explicit expression of this constant depending on the structural parameters of the system. Our argumentation is essentially based on the basic lemma describing the asymptotic expansion of the probability-generating function of the number of individuals. We state this lemma for the noncritical case. Subsequently, we find an extended analogue of the Kolmogorov constant in the noncritical case. An important role in our discussion is also played by the asymptotic properties of transition probabilities of the Q-process and their convergence to invariant measures. Obtaining the explicit form of the extended Kolmogorov constant, we refine several limit theorems of the theory of noncritical branching systems, showing explicit leading terms in the asymptotic expansions.
We study the weak convergence of the extremes of supercritical branching Lévy processes  $\{\mathbb{X}_t, t \ge0\}$ whose spatial motions are Lévy processes with regularly varying tails. The result is drastically different from the case of branching Brownian motions. We prove that, when properly renormalized,
$\{\mathbb{X}_t, t \ge0\}$ whose spatial motions are Lévy processes with regularly varying tails. The result is drastically different from the case of branching Brownian motions. We prove that, when properly renormalized,  $\mathbb{X}_t$ converges weakly. As a consequence, we obtain a limit theorem for the order statistics of
$\mathbb{X}_t$ converges weakly. As a consequence, we obtain a limit theorem for the order statistics of  $\mathbb{X}_t$.
$\mathbb{X}_t$.
Processes of random tessellations of the Euclidean space 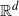 $\mathbb{R}^d$,
$\mathbb{R}^d$, 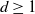 $d\geq 1$, are considered that are generated by subsequent division of their cells. Such processes are characterized by the laws of the life times of the cells until their division and by the laws for the random hyperplanes that divide the cells at the end of their life times. The STIT (STable with respect to ITerations) tessellation processes are a reference model. In the present paper a generalization concerning the life time distributions is introduced, a sufficient condition for the existence of such cell division tessellation processes is provided, and a construction is described. In particular, for the case that the random dividing hyperplanes have a Mondrian distribution—which means that all cells of the tessellations are cuboids—it is shown that the intrinsic volumes, except the Euler characteristic, can be used as the parameter for the exponential life time distribution of the cells.
$d\geq 1$, are considered that are generated by subsequent division of their cells. Such processes are characterized by the laws of the life times of the cells until their division and by the laws for the random hyperplanes that divide the cells at the end of their life times. The STIT (STable with respect to ITerations) tessellation processes are a reference model. In the present paper a generalization concerning the life time distributions is introduced, a sufficient condition for the existence of such cell division tessellation processes is provided, and a construction is described. In particular, for the case that the random dividing hyperplanes have a Mondrian distribution—which means that all cells of the tessellations are cuboids—it is shown that the intrinsic volumes, except the Euler characteristic, can be used as the parameter for the exponential life time distribution of the cells.
We study in a general graph-theoretic formulation a long-range percolation model introduced by Lamperti [27]. For various underlying digraphs, we discuss connections between this model and random exchange processes. We clarify, for all  $n \in \mathbb{N}$, under which conditions the lattices
$n \in \mathbb{N}$, under which conditions the lattices  $\mathbb{N}_0^n$ and
$\mathbb{N}_0^n$ and  $\mathbb{Z}^n$ are essentially covered in this model. Moreover, for all
$\mathbb{Z}^n$ are essentially covered in this model. Moreover, for all 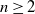 $n \geq 2$, we establish that it is impossible to cover the directed n-ary tree in our model.
$n \geq 2$, we establish that it is impossible to cover the directed n-ary tree in our model.
We consider an SIR (susceptible 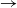 $\to$ infective
$\to$ infective 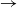 $\to$ recovered) epidemic in a closed population of size n, in which infection spreads via mixing events, comprising individuals chosen uniformly at random from the population, which occur at the points of a Poisson process. This contrasts sharply with most epidemic models, in which infection is spread purely by pairwise interaction. A sequence of epidemic processes, indexed by n, and an approximating branching process are constructed on a common probability space via embedded random walks. We show that under suitable conditions the process of infectives in the epidemic process converges almost surely to the branching process. This leads to a threshold theorem for the epidemic process, where a major outbreak is defined as one that infects at least
$\to$ recovered) epidemic in a closed population of size n, in which infection spreads via mixing events, comprising individuals chosen uniformly at random from the population, which occur at the points of a Poisson process. This contrasts sharply with most epidemic models, in which infection is spread purely by pairwise interaction. A sequence of epidemic processes, indexed by n, and an approximating branching process are constructed on a common probability space via embedded random walks. We show that under suitable conditions the process of infectives in the epidemic process converges almost surely to the branching process. This leads to a threshold theorem for the epidemic process, where a major outbreak is defined as one that infects at least 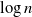 $\log n$ individuals. We show further that there exists
$\log n$ individuals. We show further that there exists 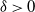 $\delta \gt 0$, depending on the model parameters, such that the probability that a major outbreak has size at least
$\delta \gt 0$, depending on the model parameters, such that the probability that a major outbreak has size at least 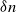 $\delta n$ tends to one as
$\delta n$ tends to one as 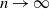 $n \to \infty$.
$n \to \infty$.
Motivated by applications to COVID dynamics, we describe a model of a branching process in a random environment 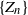 $\{Z_n\}$ whose characteristics change when crossing upper and lower thresholds. This introduces a cyclical path behavior involving periods of increase and decrease leading to supercritical and subcritical regimes. Even though the process is not Markov, we identify subsequences at random time points
$\{Z_n\}$ whose characteristics change when crossing upper and lower thresholds. This introduces a cyclical path behavior involving periods of increase and decrease leading to supercritical and subcritical regimes. Even though the process is not Markov, we identify subsequences at random time points 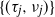 $\{(\tau_j, \nu_j)\}$—specifically the values of the process at crossing times, viz.
$\{(\tau_j, \nu_j)\}$—specifically the values of the process at crossing times, viz. 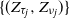 $\{(Z_{\tau_j}, Z_{\nu_j})\}$—along which the process retains the Markov structure. Under mild moment and regularity conditions, we establish that the subsequences possess a regenerative structure and prove that the limiting normal distributions of the growth rates of the process in supercritical and subcritical regimes decouple. For this reason, we establish limit theorems concerning the length of supercritical and subcritical regimes and the proportion of time the process spends in these regimes. As a byproduct of our analysis, we explicitly identify the limiting variances in terms of the functionals of the offspring distribution, threshold distribution, and environmental sequences.
$\{(Z_{\tau_j}, Z_{\nu_j})\}$—along which the process retains the Markov structure. Under mild moment and regularity conditions, we establish that the subsequences possess a regenerative structure and prove that the limiting normal distributions of the growth rates of the process in supercritical and subcritical regimes decouple. For this reason, we establish limit theorems concerning the length of supercritical and subcritical regimes and the proportion of time the process spends in these regimes. As a byproduct of our analysis, we explicitly identify the limiting variances in terms of the functionals of the offspring distribution, threshold distribution, and environmental sequences.

