
Contents
Editorial
Editorial
-
- Published online by Cambridge University Press:
- 18 September 2015, p. 5
-
- Article
-
- You have access
- Export citation
Editor’s Note
-
- Published online by Cambridge University Press:
- 18 September 2015, p. 6
-
- Article
-
- You have access
- Export citation
Research Article
The Aphasia Quotient: The Taxonomic Approach to Measurement of Aphasic Disability
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. 7-16
-
- Article
-
- You have access
- Export citation
Complex Symptomatology Simulated By Unstructured Neural Nets
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. 17-22
-
- Article
-
- You have access
- Export citation
Canadian Association of Anatomists Symposium
Functional Morphology of the Hypothalamus: Introduction
-
- Published online by Cambridge University Press:
- 18 September 2015, p. 23
-
- Article
-
- You have access
- Export citation
The Endocrine Hypothalamus: An Historical Review
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. 24-28
-
- Article
-
- You have access
- Export citation
Localization of Neuroendocrine Functions Within the Hypothalamus
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. 29-39
-
- Article
-
- You have access
- Export citation
Ultrastructure of Hypothalamic Neurons and of the Median Eminence
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. 40-58
-
- Article
-
- You have access
- Export citation
Scanning and Transmission Electron Microscopy of the Ependymal Lining of the Third Ventricle
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. 59-73
-
- Article
-
- You have access
- Export citation
Role of the Ventricular System in Neuroendocrine Processes: Synthesis and Distribution ofThyrotropin Releasing Factor (TRF) in the Hypothalamus and Third Ventricle
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. 74-84
-
- Article
-
- You have access
- Export citation
-
In vitro biosynthesis of thyrotropin releasing factor (TRF) by different regions of the hypothalamus of mink was examined. Homogenates of hypothalamic tissue were incubated in Krebs-Ringer medium containing 200 mg% glucose, 10-5M ATP, 0.1 mM histidine and glutamic acid and 0.15 μ c 3H-proline (40 Cilmmol) per mg. tissue. Extraction, purification and estimation of 3H-TRF biosynthesis involved several steps of charcoal extraction, carboxymethylcellulose and sephadex chromatography. 3H-TRF was synthesized throughout the entire antero-posterior extent of the hypothalamus in its dorsal and medial portions. 3H-TRF was synthesized also in a more discreet region, the arcuate nucleus. In vitro biosynthesis of 3H-TRF was stimulated significantly by thyroxine, but not by TSH, estradiol, corticosterone or melatonin. A method is described for collection of cerebrospinal fluid of the third ventricle of the rat brain; TRF concentration in this fluid was approximately
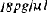 in normal animals.
in normal animals.The distribution of TRF-producing cells in the hypothalamus and presence of TRF in cerebrospinal fluid of the third ventricle is discussed with respect to the hypothesis that this releasing factor may be delivered to the median eminence and adenohypophysis in part, via the cerebrospinal fluid.
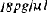 in normal animals.
in normal animals.Announcement
Announcements
-
- Published online by Cambridge University Press:
- 18 September 2015, p. 85
-
- Article
-
- You have access
- Export citation
Front Matter
CJN volume 1 issue 1 Cover and Front matter
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. f1-f6
-
- Article
-
- You have access
- Export citation
Back Matter
CJN volume 1 issue 1 Cover and Back matter
-
- Published online by Cambridge University Press:
- 18 September 2015, pp. b1-b3
-
- Article
-
- You have access
- Export citation